|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
СТРОЕНИЕ КОРЫ ГОЛОВНОГО МОЗГА И ПРОВОДЯЩИЕ ПУТИ ГОЛОВНОГО МОЗГАЦитоархитектонически во вторичных полях ядерных зон коры выступают на первый план те элементы их нейронной структуры, которые приспособлены для переключения приходящих из подкорки афферентных импульсов через посредство зерновидных клеток слоя IV на крупные пирамиды нижнего
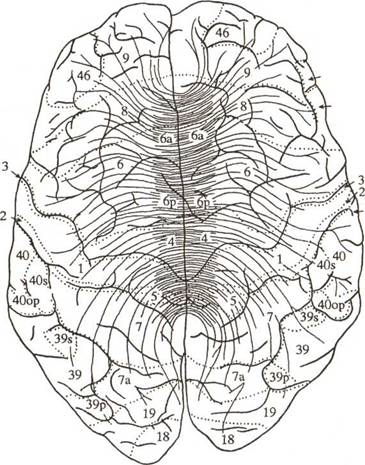
Рис. 6. Топографическая карта мозговой коры (по Г.И. Полякову). Ядерные и внеядерные зоны коры головного мозга — наружная поверхность. Ядерные зоны мозговой коры обозначены кружками (зрительная зона), квадратами (слуховая зона), ромбами (общечув^ви-тельная зона), треугольниками (двигательная зона); центральные поля выделены крупными знаками. Зоны перекрытия анализаторов ij задних отделах полушария (теменно-височно-затылочные и нижнете/Мен-ные отделы) обозначены смешанными знаками; в передних отделах полушария (лобная область) — видоизмененными треугольниками; лимбическая и инсуларная области, а также филогенетически старые зоны коры — прерывистой штриховкой.
Вторичные поля ядерных зон (рис. 6, 7, 8, 9) играют весьма существенную роль в осуществлении взаимной связи отдельных раздражителей, выделяемых через посредство первичных полей, а также в функциональном объединении ядерных зон разных анализаторов в процессе интеграции сложных комплексов рецепций, идущих по разным анализаторам. Эти поля имеют, таким образом, преимущественное отношение к осуществлению более сложных форм координированных психических процессов, связанных с детальным анализом соотношений конкретных предметных раздражителей и с ориентировкой в конкретном окружающем пространстве и времени. Возбуждение, возникающее во вторичных полях, имеет тенденцию распространяться на значительно более широкие территории коры за пределами данной ядерной зоны.  Афферентные импульсы, направляющиеся в кору от рецеп-торных поверхностей органов чувств, достигают этих полей через большое число дополнительных переключений в так называемых ассоциационных ядрах зрительного бугра в отличие от афферентных импульсов, направляющихся в первичные поля коры более коротким путем через так называемые реле-ядра зрительного бугра (А. Р. Лурия, 2000, стр. 47). «Морфологическим выражением дальнейшего усложнения функциональных отношений между анализаторами является тот факт, что в процессе прогрессивного роста ядерных зон анализаторов по поверхности полушария возникает взаимное перекрытие этих зон с образованием особых «зон перекрытия» корковых концов анализаторов. Эти корковые формации, представленные в задней части полушария полями верхней и нижней теменной областей, средней височной и височно-те-менно-затылочной подобластей, имеют отношение к наиболее сложным формам интеграции совместной деятельности зрительного, слухового и кожно-кинестетического анализаторов. Как мы увидим ниже, при поражении этих территорий расстраиваются главным образом наиболее обобщенные проявления корковой деятельности, опирающиеся на совместную работу нескольких анализаторов, позволяющие осуществлять наиболее сложные формы ориентировки во внешнем мире и анализ и синтез сложных систем отношений между раздражителями, воздействующими на воспринимающую часть разных анализаторов» (А.Р. Лурия, там же). Богатый клинический опыт учит, однако, что эти «рассеянные» элементы анализаторов в максимальной степени концентрируют в себе потенциальные возможности функциональных взаимодействий между анализаторами, протекающих притом на высшем уровне. Обширная у человека территория, занимаемая зонами перекрытия корковых концов анализаторов, распадается на ряд полей. Особенности микроструктуры этих полей, так же как и их функциональное значение, определяются топографическими соотношениями этих полей с полями ядерных зон, между которыми они располагаются. Оправданным представляется выделение трех частных зон перекрытия, представленных соответственно верхней теменной областью, нижней теменной областью и височно-теменно-затылочной подобластью (А.Р. Лурия, там же). Обе теменные области топографически и функционально наиболее тесно связаны с зонами кожно-кинестетического и зрительного анализаторов, между которыми эти области располагаются. Верхняя теменная область граничит спереди с теми разделами постцентральной области, которые заняты представительством ног и туловища. Эта область соответственно имеет специальное значение для интеграции со зрительными рецепциями движений всего тела. Нижняя теменная область граничит с теми разделами постцентральной области, которые заняты представительством рук и лица. Соответственно та область имеет отношение к интеграции обобщенных и отвлеченных форм сигнализации, которые связаны с тонко и сложно дифференцированными предметными и речевыми действиями, совершаемыми под контролем зрения и требующими совершенно разработанной системы ориентировки в окружающем пространстве (рис. 9). Наконец, височно-теменно-затылочная подобласть, составляющая зону перехода между слуховой и зрительной зонами коры, как показывают клинические наблюдения, функционально особенно тесно связана с наиболее сложными формами интеграции слуховых и зрительных рецепций. Наряду с весьма дробно дифференцированными многосторонними ассоциационными связями третичные поля коры характеризуются по сравнению с первичными и вторичными полями и наиболее сложными цепями проекционных переключений в подкорковых отделах анализаторов (рис. 8, 9).
Рис. 9. Схема соматотопической проекции в коре головного мозга (по Пенфилду). А — корковая проекция чувствительности; Б — корковая проекция двигательной системы. Относительные размеры органов отражают ту площадь коры головного мозга, с которой могут быть вызваны соответствующие ощущения и движения.
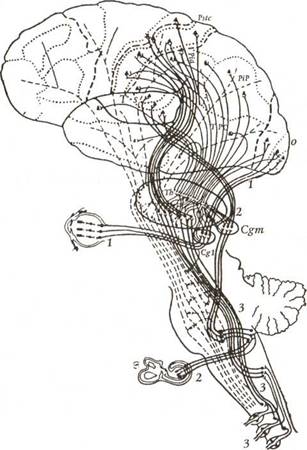
Рис. 10. Схема корково-подкорковых соотношений первичных, вторичных и третичных зон (по Г.И. Полякову). Жирными линиями показаны системы анализаторов с их переключениями в подкорковых отделах: 1 — зрительный анализатор; 2 — слуховой анализатор; 3 — кожно-кинестетический анализатор. Т — височная область; О — затылочная область; Pip — поле 39; Pia — поле 40; Psts — постцентральная область; ТРО — височно-теменно-затылочная подобласть; Th — зрительный бугор; Cgm — внутреннее коленчатое тело; Cgl — наружное коленчатое тело.
Суммируя все вышесказанное, пишет А.Р. Лурия, следует особенно подчеркнуть, что в нормально действующем мозге описанные выше три группы корковых полей вместе с их системами переключений и связей между собой и с подкорковыми инстанциями анализаторов работают как одно сложнодиф-ференцированное целое. В онтогенезе человека этот период падает на первые недели и месяцы жизни. Соответственно и процесс миэлинизации проводящих путей, связанных с этими полями, протекает в более поздние сроки. Наиболее поздно созревают зоны перекрытия анализаторов и формации лобной области; это наблюдается только на протяжении первых лет жизни ребенка. Связи этих корковых территорий также наиболее поздно завершают цикл своего развития. Специфически человеческой особенностью являются существенные преобразования корковой организации, вызванные появлением второй сигнальной системы, связанной с трудовой деятельностью человека. Г.И. Поляков, излагая проблемы структуры движения, пишет, что взаимодействие составляющих, определяющих фактическое перемещение в пространстве отдельных звеньев кинематической цепи, определяется следующими тремя основными факторами: а) физиологическими состояниями самих мышц; б) влиянием внешних сил (тяжести, инерции, ускорения, сопротивления объекта движения или окружающей среды); в) равнодействующей реактивных сил или сил отдачи, которые быстро возрастают с увеличением числа звеньев цепи и степеней свободы подвижности. Соответственно имеется много возможных траекторий подлежащего осуществлению движения, то есть проекций его в определенной системе пространственных координат в определенные отрезки времени. В процессе реализации движения важное значение имеют эффекты установки («преднастройки») элементов рефлекторного прибора на очередную, предстоящую фазу его деятельности. Реальное движение строится, таким образом, на непрерывной функциональной связи отдельных его компонентов благодаря каждомоментному сличению текущего отрезка движения со свежим следом от предшествующего; одновременно с этим происходит опережение в микроинтервалах времени осуществляемого в данный момент двигательного акта и установка его на последующий отрезок движения (феномен «опережающей преднастройки», по Н.А. Бернштейну). Нарушения коррекционной обратной связи особенно демонстративно выявляются при заболеваниях мозжечка, который представляет собой орган, приспособленный для осуществления бессознательно рефлекторным, автоматическим грутем самоконтроля над всеми движениями и положениями тела в пространстве. «Сложным образом проявляется, — пишет Г.И. Поляков, — участие обратной сигнализации и в функции устной речи, которая строится на физиологической основе взаимодействия высокодифференцированных и весьма тонко специализированных кинестетических и слуховых рецепций. Произносимое слово должно соответствовать слову слышцМому. Движения и положения различных частей речевого аппарата при производстве членораздельных звуков слов не тольк^ регулируются кинестетическими сигналами, поступающими; непосредственно от самого речевого аппарата (голосовых связок, мышц губ и языка), но и контролируются через посредство воспринимаемых на слух звуков речи. Слуховой контроль является необходимым этапом в процессе овладения устной речью ребенком, а также при обучении взрослого иностранному языку. Значение обратной сигнализации через слуховой анализатор для деятельности речевого аппарата выступает особенно наглядно, если принять во внимание речевые расстройства, наступающие при дефектах данной сигнализации у глухих от рождения, устная речь которых часто отличается невнятностью и мал ©выразительностью». Следует подчеркнуть, что условный рефлекс образуется лишь на те сигналы, которые так или иначе подкрепляется, то есть сопровождаются определенной обратной сигнализацией. Никак не подкрепляемые сигналы не могут служить физиологической базой для установления временной связи.
Рис. 11. Схема цитоархитектонических полей и ассоциационных путей большого мозга человека. Наружная поверхность полушария мозга с изображением хода дугообразных волокон, верхнего продольного и крючковидного пучков и обозначением цитоархитектонических полей.
Рис. 12. Схема комиссуральных путей и цитоархитектонических полей большого мозга человека (по СБ. Дзугаевой). Дорзальная поверхность полушарий мозга с изображением хода волокон мозолистого тела и обозначением цитоархитектонических полей.
Кроме исследований А.Р. Лурия, Г.И. Полякова и ряда невропатологов, нельзя не сослаться на исследования СБ. Дзугаевой, акцентирующей наше внимание на ассоциативных связях лобной, височной, затылочной и теменной долей (рис. 11, 12 см. на форзацах). СБ. Дзугаева (1975) прослеживает весь ход развития ассоциативных путей с момента рождения человека до зрелого возраста. В связи с важностью ее исследований приведем эти сведения. Ассоциативные пути формируются одновременно с развитием полей коры конечного мозга в тесной связи с другими системами. В процессе дифференцировки определяются топографические соотношения, намечаются положение и форма отдельных пучков, принимающих участие в формировании ассоциативной системы. Одни группы волокон принимают более стабильные, другие — более пластические формы в онтогенезе. В связи с этим их можно разделить на две группы: в первой форме ассоциативные пути резко не изменяются в течение всех этапов развития, а сохраняются более или менее устойчиво; во второй форме меняются в сторону установления более тесных связей с неокортикальными формациями. Отмечается неодновременное развитие отдельных пучков ассоциационной системы. Различные темпы формирования и созревания отдельных пучков одной и той же системы связаны с гетерохронным ростом сложной по строению и функции коры большого мозга человека. Ассоциационные пути, являясь анатомическим субстратом сочетательной деятельности, обусловливают взаимодействие корковых концов анализаторов и интегративную функцию больших полушарий мозга. Они обеспечивают нарастающее в онтогенезе совершенствование рефлекторных механизмов конечного мозга. Чем сильнее развиты ассоциационные пути, тем шире и многообразнее представлена взаимосвязь между корковыми зонами зрительного, слухового, двигательного, вкусового, обонятельного и других анализаторов. Дифференцировка ассоциационных путей на анатомически обособленные пучки нервных волокон становится макроскопически видимой во второй половине пренатального онтогенеза. Характерные особенности хода каждого пучка выявляются уже к моменту рождения. В постнатальном онтогенезе продолжается дальнейший прогрессивный рост всех ассоциационных путей — постепенно они приобретают компактность, извилистость, изменяются объемные соотношения между отдельными
Рис. 13. Мозг взрослого человека. Проводящие пути (по СБ. Дзугаевой). 1 — верхний продольный пучок — fasciculus longitudinals superior; 2 — крючковидный пучок — fasciculus incinatus; 3 — наружная сумка — capsula externa; 4 — лобная доля — lobus frontalis; 5 — затылочная доля — lobus occipitalis; 6 — височная доля — lobus temporalis; 7 — теменная доля — lobus parietalis.
Рис. 14. Мозг взрослого человека. Проводящие пути (по СБ. Дзугаевой). 1 — верхний продольный пучок — fasciculus longitudinals superior; 2 —крючковидный пучок — fasciculus incinatus; 3 — наружная сумка — cap
ассоциационными пучками, увеличивается зона их распределения в полях коры большого мозга. В более поверхностных слоях больших полушарий располагаются короткие ассоциационные волокна, а в более глубоких слоях — длинные. В составе поясного, крючковидного, нижнего и верхнего продольного пучков содержатся короткие и длинные волокна; первые лежат поверхностно и имеют дугообразно-извилистое направление, вторые расположены глубже и принимают более прямолинейный ход. Послойное изучение ассоциационных путей в онтогенезе человека показывает, что длина их увеличивается с поверхностных отделов вглубь. Процесс развития ассоциационных путей идет в такой последовательности: вначале формируются пучки, связанные с корковым концом обонятельного и вкусового, затем слухового и зрительного анализаторов. Чем сильнее развиты борозды и извилины, тем более выражена извилисто-волнообразная форма ассоциационных путей. Сравнительная характеристика их на разных этапах постна-тального онтогенеза говорит о том, что эти пути подразделяются на широкие и узкие, короткие и длинные. Обозначение «широкие и узкие» получает различное значение в разные периоды развития. Так, верхний продольный пучок у плода имеет широкую форму, у детей — узкую, а у взрослых — опять широкую. Онтогенетические данные, о которых идет речь в настоящей главе, говорят об известном разнообразии и характерных особенностях в общем плане структурной организации ассоциационной системы. Ассоциационные пути характеризуются тем, что они широко распространяются в полях коры большого мозга, контактируют непосредственно с корковыми концами всех анализаторных систем, обеспечивая, таким образом, их сочетательную деятельность на корковом уровне. В процессе развития ассоциационных путей увеличиваются их число, размеры, объемные соотношения, степень миелинизации. Топографо-анатомические взаимоотношения проводящих путей мозга с возрастом усложняются, увеличивается взаимосвязь между ассоциационными, комиссуральными и проекционными путями. Ассоциационные пути приобретают довольно обширную корковую область распределения. Они устанавливают взаимосвязь между функционально различными полями коры большого мозга, благодаря чему создаются обширные возможности для условно-рефлекторной и сочетательной деятельности конечного мозга. Усложнение структурной организации ассоциационных путей в процессе онтогенетического развития свидетельствует о все возрастающей роли их у детей старшего возраста и у взрослых. В онтогенезе, как и в филогенезе, изменяются соотношения между проекционными и ассоциационными путями в сторону преобладания последних; это является показателем того, что с возрастом филогенетически новые пути начинают преобладать над филогенетически старыми. Интенсивное развитие полей коры, центральных звеньев анализаторов влияет направляющим образом на формирование ассоциационных и комиссуральных путей, делает их более дифференцированными и тонко организованными у человека, что имеет важное значение для функциональной взаимосвязи между анализаторами на неокортикальном уровне. Как видно из вышеприведенных рисунков (1, 2, 4, 6, 7, 8, 9 и др.), все ядерные первичные и вторичные поля речевых зон сосредоточены вблизи друг от друга (поля 44, 45; 1, 7, 40, 39, 22, 21), в местах прохождения наиболее крупных передних и задних ветвей левой средней мозговой артерии, вдоль которых располагаются связывающие их проводящие пути. Разнообразие и сложность анатомических путей, обеспечивающих взаимодействие и распространение информации различной модальности на корковом уровне, являются морфологической основой компенсаторно-приспособительных возможностей нервной системы. С усложнением межполушарных и внутриполушарных взаимоотношений расширяется мультисенсорное взаимодействие афферентных систем на корковом уровне, что обеспечивает совершенствование центральных механизмов целостных системных реакций растущего организма ребенка. Ассоциационные и комиссуральные пути устанавливают структурную и функциональную взаимосвязь между близлежащими и отдаленными точками коры, благодаря чему сложнодифференци-рованные корковые структуры объединяются в один координационный механизм для гармоничной деятельности мозга в целом. Процесс усложнения в онтогенезе структурно-функциональной организации проводящих путей в системе анализаторов соответствует развитию и усложнению полей коры конечного мозга. В онтогенезе в связи с прогрессивной дифференцировкой и развитием неокортекса меняются топографические взаимоотношения между проводящими путями мозгового ствола и конечного мозга. Данные ассоциативные связи мозга приведены и в работах А.Р. Лурия (1966). Так, А.Р. Лурия обращает специальное внимание на взаимодействие префронтальной (заднелобной) области с затылочной, височной и теменной долями. Все эти ценнейшие сведения об огромной системе проводящих путей чрезвычайно важны при понимании компенсаторных процессов преодоления нарушений ВПФ при сосудистых поражениях головного мозга.
|
|||||||
|
|
Последнее изменение этой страницы: 2018-05-10; просмотров: 512. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |