|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
Неупорядоченная вторичная структура (конформация клубка)
Те участки белковой молекулы, которые не относятся ни к спирализованным, ни к складчатым структурам, обычно называют неупорядоченными. В конформации клубка может находиться значительная часть белковой молекулы. Нужно отметить, что с точки зрения биологической значимости неупорядоченные участки белка столь же важны, как a-спираль и b-структура.
Третичная структура
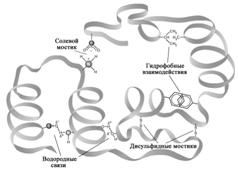
Третичной структурой белковых молекул называется трехмерная пространственная организация белка, формирующаяся и стабилизирующаяся за счет многих и различных по природе слабых взаимодействий между остатками аминокислот, которые могут располагаться на значительном расстоянии друг от друга в составе полипептидной цепи. К таким взаимодействиям относятся солевые мостики, водородные связи, гидрофобные взаимодействия, а также ковалентные дисульфидные связи (Рис. 2.6).
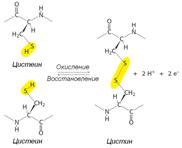
Рисунок 2.6 – Связи и взаимодействия в третичной структуре белка. Гидрофобные взаимодействия. Неполярные молекулы или части молекул не способные взаимодействовать с водой называются гидрофобными. К гидрофобным группам относятся алкильные и арильные (ароматические) радикалы ряда аминокислот. Гидрофобные радикалы аминокислотных остатков избегают контактов с водным окружением и стремятся собраться вместе внутри глобулярной структуры, где они формируют гидрофобное ядро. Электростатические взаимодействия.К ним относятся ион-ионные, ион-дипольные и диполь-дипольные взаимодействия. Эти ионные связи возникают между разноименно заряженными группами боковых цепей аминокислот. Например, при физиологических значениях рН e-аминогруппа лизина несет заряд +1, а карбоксильная группа глутаминовой кислоты в составе боковой цепи несет заряд -1. Следовательно, боковые радикалы указанных аминокислот способны взаимодействовать друг с другом электростатически, дополнительно стабилизируя структуру белка, за счет образования солевых мостиков.  Дисульфидные мостики.Третичная структура ряда белков может стабилизироваться дисульфидными мостиками (—S— S—), образующимися за счет ковалентного взаимодействия SH-групп двух остатков цистеина (Рис. 2.7). Принимающие участие в образовании (—S—S—)-мостиков остатки цистеинов могут быть линейно существенно разобщены в полипептидной цепи, однако при формировании третичной структуры SH-группы таких остатков цистеина оказываются пространственно сближенными и ковалентно сшивают два участка одной полипептидной цепи (или соседних цепей).
Рисунок 2.7 – Механизм образования S—S-мостиков между SH-группами остатков цистеина линейно разобщенными в полипептидной цепи, но пространственно сближенными.
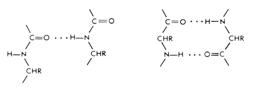
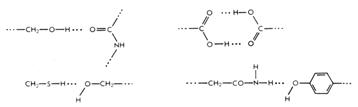
Водородные связи.В отличие от водородных связей поддерживающих вторичные структуры белков, водородные связи, участвующие в стабилизации третичной структуры более разнообразны (Рис. 2.8). В этом случае водородные связи образуются: - между боковыми R-группами аминокислот; - между карбонильными атомами кислорода и атомами амидного водорода, принадлежащими пептидным группам остова; - между поверхностными остатками полярных аминокислот белка и молекулами воды, расположенными в соседних петлях полипептидной цепи; - между, например, гидроксильной группой остатка серина в одном участке полипептидной цепи и атомом азота в кольце остатка гистидина, находящегося в соседней петле той же цепи; - между амидной группой глутамина в одном участке полипептидной цепи и остатком тирозина в другом участке.
Рисунок 2.8 – Различные варианты формирования водородных связей в третичной структуре белка.
Четвертичная структура
Четвертичной (олигомерной) структурой обладают белки, построенные из двух и более полипептидных цепей, каждая из которых уложена в соответствующую третичную структуру. Олигомерная организация белка стабилизируется водородными связями, электростатическими и гидрофобными взаимодействиями между аминокислотными остатками, находящимися на поверхности индивидуальных полипептидных цепей. Полипептидные цепи, соответствующие каждому из олигомеров белка получили название мономеров или субъединиц. Субъединицы, формирующие четвертичную структуру олигомерного белка, могут быть одинаковыми или разными, другими словами они могут иметь одинаковую молекулярную массу и обладать одинаковой третичной организацией, а могут отличаться друг от друга по указанным параметрам. В тоже время под протомером понимают наименьшую повторяющуюся часть олигомера. Многие олигомерные белки состоят из двух или четырех субъединиц и называются, соответственно, димерами или тетрамерами. Однако довольно часто встречаются олигомеры, содержащие более четырех субъединиц. Это особенно характерно для регуляторных белков.
Рисунок 2.9 – Принцип формирования гомоолигомеров.
Олигомерные белки играют особую роль во внутриклеточной регуляции: их протомеры способны циклически (т.е. обратимо) изменять взаимную ориентацию, что приводит к изменению свойств всего олигомера. Наиболее изученным примером олигомерного белка является гемоглобин. Роль первичной структуры в формировании более высоких уровней структурной организации белков трудно переоценить. Действительно, вторичная и третичная структуры белков, формирующиеся самопроизвольно, определяются последовательностью аминокислот в полипептидных цепях. Эти процессы детерминируются химическими группами, соединенными с a-углеродными атомами аминокислотных остатков. Таким образом, можно сказать, что не существует независимого генетического контроля по формированию уровней структурной организации белков выше первичного, поскольку первичная структура специфически определяет и вторичную, и третичную, и четвертичную структуры. Нативной конформацией белка, по-видимому, следует считать термодинамически наиболее устойчивую структуру в конкретных условиях.
|
||||
|
|
Последнее изменение этой страницы: 2018-04-12; просмотров: 432. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |