|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
Экзаменационный билет № 68.ОБЩАЯ МИКРОБИОЛОГИЯ Экология вирусов-раздел экологической микробиологии, исследующий взаимодействие вирусов между собой и с экологическими факторами среды обитания. Экология вирусов изучает экологические среды, формы и факторы эволюционной изменчивости, потоки движения генетического материала в популяциях, закономерности формирования видовых генофондов, влияние физ. и хим. факторов на вирусы, способы и методы использования экологических знаний в борьбе с вирусными инфекциями. ПЕРВЫМ методом является заражение лабораторных животных. После стали использоваться куриные имбрионы для репродукции вирусов с диагностич целями. Поскольку основу всего живого составляют генетические структуры, то и вирусы КЛАССИФИЦИРУЮТ сейчас по характеристике их наследственного вещества - нуклеиновых кислот. Все вирусы подразделяют на две большие группы: ДНК-содержащие вирусы (дезоксивирусы) и РНК-содержащие вирусы (рибовирусы). ДНК-содержвирусы им ДНКгеном,транскрибируются в ядре клетки с пом клеточной РНК –полимеразы,в рез чего обра-ся ирНК,которая транслируется с образование белков вируса. Особеннотьэтого процесса явл-ся синтез иРНК в ядре с пом клеточной РНК-полимеразы(аденовирусы,ВПЧ) или с пом собственной РНК-полимеразы(поксивирусы). Они бывают однонитевым(на исходной вирусной ДНК (плюс нить) комплементарно синтезируется минус нить ДНК, служащая матрицей в синтезе плюс нити ДНК для новых поколений вирусов. Параллельно синтезируется иРНК, происходит трансляция вирусных белков, которые возвращаются в ядро, где собираются вирионы) и двунитевыми(герпес) Репликация двунитевых вирусных ДНК проходит обычным полуконсервативным механизмом: после расплетения нитей ДНК к ним комплементарно достраиваются новые нити  Плюс-нитевые РНК сод вирусы(пикорнавирусы) им геном выполн ф-ии иРНК.Белки этих вирусов синт-ся в клетке без транскрипции Минус-нитевые РНКвирусы(ортомикровирусы) им геном,выполн роль матрицы.с котрой транскрибируется иРНК, при уч РНК-полимеразы Ретровирусы(ВИЧ) имеют диплоидный геом,сост из двух идентичных РНК. Они имеют обратную транскриптазу, с пом кот осущ-т обратную транскрипцию(синтез нити ДНК) Комплементарная нить копир-ся с образо-м двунитевой ДНК,кот встраив-ся в клеточный геном и в его составе транскрибируется в иРНК ИММУНОЛОГИЯ 3,4
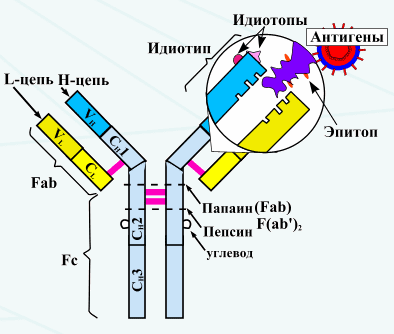
Антитела- специфические эффекторы гуморального иммунитета. Биохим и клеточная природа антител: 1)крупные гликопротеины 2)белки плазмы крови, участвующие в ИО-иммуноглобулины 3) при электрофорезе АТ мигрируют в основном с фракцией гаммаглобулинов Клеточная природа антител: Продуцируются плазматическими кл- потомками клона специфических В-лимфоцитов, котрые контактировали с соотв антигеном Специфичность АТ: конкретный Ig способен связываться с АГ, вызвавшим его образование. Антитела имеют форму буквы Y: каждая структурная единица-домен; с антигеном взаимодействуют вариабельные домены. Большинство молекул иммуноглобулинов составлено из двух тяжелых (H) цепей и двух легких (L) цепей, соединенных дисульфидными связями. Легкие цепи состоят или из двух k-цепей, или из двух l-цепей. Тяжелые цепи могут быть одного из пяти классов (IgA, IgG, IgM, IgD и IgE).
Каждая цепь имеет два участка: 1) постоянный. Остается постоянным в последовательности аминокислот и антигенности в пределах данного класса иммуноглобулинов; 2) вариабельный. Характеризуется большой непостоянностью последовательности аминокислот; в этой части цепи происходит реакция соединения с антигеном. Каждая молекула IgG состоит из двух соединенных цепей, концы которых формируют два антигенсвязывающих участка. На вариабельном участке каждой цепи имеются гипервариабельные участки: три в легких цепях и четыре в тяжелых. Разновидности последовательности аминокислот в этих гипервариабельных участках определяют специфичность антитела. При определенных условиях эти гипервариабельные области могут также выступать в роли антигенов (идиотипов). Функции антител: 1) Первичная: специфическое связывание Аг 2) Вторичные : · Активация системы комплемента · Обсоническая функция · Прикрепление к Fc рецепторам клеточных мемран · Участвуют в переносе IgG через плаценту
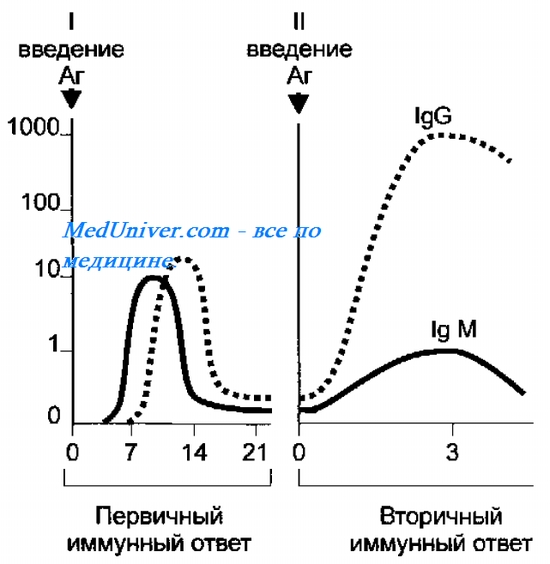
Понятия об аллотипах, идиотипах. Аллотипантител определяется константными доменами и каркасными участками вариабельных доменов H и L цепей. Идиотипопред-ся структурным своеобразием вариабельных доменов, формирующих паратоп. Динамика антител в ходе первичного и вторичного ИО: После поступления чужеродного АГ в организм через несколько дней начинают появляться антитела плазмы крови. В процессе инфекционного заболевания концентрация антител увеличивается. Приблизительно через 3 дня появляются IgM, затем в небольшом кол-ве IgG. Реакция на первый и повторный контакт отличаются.
Функции антител: 3) Первичная: специфическое связывание Аг 4) Вторичные : · Активация системы комплемента · Обсоническая функция · Прикрепление к Fc рецепторам клеточных мемран · Участвуют в переносе IgG через плаценту
БАКТЕРИОЛОГИЯ Таксономия: порядок Chlamydiales, семейство Chlamydaceae, род Chlamydia. Род представлен видами С.trachomatis, C.psittaci,C.pneumoniae.
Хламидии (Chlamydia) -- мелкие грамотрицательные кокковидные бактерии, размером 250-1500нм (0,25-1 мкм). Они имеют все основные признаки бактерий: содержат два типа нуклеиновых кислот (ДНК и РНК), рибосомы, мурамовую кислоту (компонент клеточной стенки грамотрицательных бактерий), размножаются бинарным делением и чувствительны к некоторым антибиотикам. Хламидии неспособны самостоятельно синтезировать высокоэнергетические соединения и обеспечивать собственные потребности в энергии. Нормальное развитие хламидий возможно только в условиях внутриклеточного паразитирования.
Хламидии размножаются бинарным делением. Жизненный цикл хламидии включает образование двух основных форм (рис. 23-1). • Элементарное тельце хламидии (ЭТ) — мелкая (0,2-0,5 мкм) сферическая внеклеточная структура с трёхслойной клеточной стенкой. Метаболически малоактивно и адаптировано к внеклеточному выживанию. Элементарные тельца хламидий — инфекционные единицы, заражающие клетки. По Романовскому-Гимзе ЭТ окрашиваются в пурпурный цвет. • Ретикулярное тельце хламидии (РТ) — репродукционная внутриклеточная форма. Представлено более крупным образованием (до 1 мкм), имеющим сетчатую структуру с тонкой клеточной стенкой. Развивается в течение 5-6 ч из ЭТ, проникшего в цитоплазму и претерпевшего структурные изменения. Первоначально из ЭТ образуется инициальное тельце (вегетативная форма), окрашивающееся в синий цвет по Романовскому-Гимзе. Затем инициальное тельце превращается в РТ. ВИРУСОЛОГИЯ Структура ВИЧ относится к семейству ретровирусов. Вирион имеет сферическую форму, диаметром 100–150 нм. Кубический тип симметрии. Наружная (суперкапсидная) оболочка вируса состоит из бимолекулярного слоя липидов, который имеет происхождение из клеточной мембраны клетки хозяина. Из нее выступают шипы двух типов: 1) gp 120 (обладает рецепторной функцией); 2) gp 41 (обладает якорной функцией). В эту мембрану встроены рецепторные образования. Под наружной оболочкой располагается сердцевина вируса (кор), которая имеет форму усеченного конуса. Промежуток между наружной вирусной мембраной и сердцевиной вируса заполнен матриксным белком. Внутри сердцевины располагаются две одинаковые молекулы вирусной РНК, связанные с низкомолекулярными белками р6 и р7. Каждая молекула РНК содержит девять генов ВИЧ: 1) структурные (три гена); 2) регуляторные (три гена, они не кодируют структурных компонентов вируса, но, попав в клетку, кодируют образование веществ, которые либо угнетают активность структурных генов, либо активируют); 3) дополнительные (три гена, они содержат информацию, необходимую для продукции белков, которые управляют способностью вируса инфицировать клетку, реплицироваться и вызывать заболевание). Выделяют три группы структурных генов: 1) gag (кодируют образование структурных белков сердцевины вируса); 2) pol (направляют синтез белков – вирусных ферментов); 3) ent (кодируют синтез оболочечных белков gp 120 и gp 41). Концы каждой молекулы РНК содержат дублированную последовательность РНК. Эти участки действуют как переключатели для управления процессом вирусной транскрипции, взаимодействуя с белками ВИЧ или белками клетки хозяина. Кроме РНК, там же находятся вирусные ферменты: 1) обратная транскриптаза; осуществляет синтез вирусной ДНК с молекулы вирусной РНК; 2) протеаза; участвует в «нарезании» предшественников вирусных белков при созревании новой вирусной частицы; 3) эндонуклеаза (интеграза); производит встраивание вирусной ДНК в геном клетки хозяина, в результате чего образуется провирус. Антигенными свойствами обладают: 1) белки сердцевины; 2) оболочечные гликопротеины. Характеризуются высоким уровнем антигенной изменчивости, который определяется высокой скоростью замен нуклеотидов. Интенсивная антигенная изменчивость ВИЧ происходит в организме больных в ходе инфекции и у вирусоносителей. Она дает возможность вирусу «скрыться» от специфических антител и факторов клеточного иммунитета, что приводит к хронизации инфекции. В обычных культурах клеток ВИЧ не культивируется. Для культивирования используется культура Т-лимфоцитов с хелперной функцией. |
||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2018-04-12; просмотров: 402. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |