|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
ПРОВОДЯЩИЕ ПУТИ ГОЛОВНОГО И СПИННОГО МОЗГА
Проводящие пути являются частью сложных рефлекторных дуг, которые соединяют между собой различные отделы центральной нервной системы и обеспечивают двухстороннюю функциональную связь между отдельными структурами головного и спинного мозга. Они отличаются многочисленностью, сложностью строения и надежностью функционирования. В зависимости от величины, формы и направления нервного импульса проводящие пути получают название: путь (tractus), пучок (fasciculus), волокна (fibrae), спайка (commissure), петля (lemniscus) или лучистость (radicitio). Все проводящие пути ЦНС подразделяют на три группы: 1. Проекционные. 2. Комиссуральные. 3. Ассоциативные. В процессе становления проводящей системы в онтогенезе первоначально формируются проекционные пути, а затем комиссуральные и ассоциативные.
ПРОЕКЦИОННЫЕ ПРОВОДЯЩИЕ ПУТИ
Проекционные проводящие пути, neurofibraeprojectiones, связывают кору головного мозга с его нижележащими отделами (короткие проводящие пути) и со спинным мозгом (длинные проводящие пути). Но направлению проведения нервного импульса проекционные пути подразделяют на две группы: • афферентные (восходящие, центростремительные, чувствительные), которые проводят нервный импульс от рецепторов, воспринимающих информацию из внешнего мира или внутренней среды организма к различным отделам головного мозга и к коре полушарий; • афферентные (нисходящие,центобежные, двигательные), передающие импульс от коры головного мозга и других его отделов на периферию.
ЧУВСТВИТЕЛЬНЫЕ ПРОВОДЯЩИЕ ПУТИ
Под чувствительностью понимают способность организма воспринимать воздействия раздражителей внешней и внутренней среды.  Различают следующие виды чувствительности: 1. Поверхностная (кожная), или экстероцеитивная чувствительность — тактильная, болевая, температурная. 2. Глубокая, или проприоцептивная чувствительность — мышечно-суставная, вибрационная, чувство давления и веса. 3. Иитероценгивная, или вегеговисцеральная чувствительность — чувствительность внутренних органов и сосудов. Вид чувствительности определяется характером рецептора, воспринимающего раздражение. Среди рецепторов различают экстерореценторы, располагающиеся в коже и слизистых оболочках; проприорецепторм. которые находятся в мышцах, сухожилиях, связках, суставных капсулах, и висцерорецепторм, расположенные в различных внутренних органах и сосудах. В зависимости от вида чувствительности афферентные проводящие пути подразделяют на пути: • экстероцептивной, • проприоцептивной, • интероцептивной чувствительности. Те из них, которые заканчиваются в коре полушарий большого мозга, получили название проводников сознательной чувствительности, или проводящих путей коркового направления. ЧУВСТВИТЕЛЬНЫЕ ПРОВОДЯЩИЕ ПУТИ КОРКОВОГО НАПРАВЛЕНИЯ
Первые (рецепторные) нейроны чувствительных проводящих путей вынесены за пределы головного и спинного мозга в чувствительные ганглии. Это спинномозговые узлы или чувствительные ганглии черепных нервов. Первые нейроны чувствительных путей представлены биполярными (псевдоуниполярными) клетками. Их дендриты (периферические отростки) в составе спинномозговых или черепных нервов направляются на периферию и заканчиваются рецепторами. Рецепторы воспринимают раздражение из внешней или внутренней среды, преобразуя его энергию в нервный импульс, который по дендриту передается к телу нервной клетки, а затем — к ее аксону. Аксоны (центральные отростки) в составе задних корешков спинномозговых или чувствительных корешков черепных нервов входят в вещество мозга и разделяются на пучки в зависимости от проводимой ими чувствительности. В спинном мозге афферентные волокна занимают строго определенное положение (рис. 3.1). Далее аксоны достигают соответствующих ядер спинного мозга или мозгового ствола, где заканчиваются на телах вторых нейронов. В области моста и среднего мозга аксоны вторых нейронов поднимаются в составе так называемых чувствительных петель (медиальной, спинномозговой и тройничной), которые заканчиваются в ядрах латеральной группы зрительного бугра на телах третьих нейронов. Аксоны третьих нейронов проходят через заднюю ножку внутренней капсулы (рис. 3.2) и направляются в соответствующие центры коры полушарий большого мозга (интеграционные центры), образуя в белом веществе полушарий вместе с волокнами двигательных путей лучистый венец. Таким образом, все афферентные пути, проводящие разнообразные чувствительные импульсы, переключаются в таламусе, который является местом интеграции сенсорной информации.
Рис. 3.1. Топография серого и белого вещества сшитого мозга: 1 — rr. tectospinalis; 2 — tr. corticospinalis anterior; 3 — tr. spinotlwlamicus anterior; 4 — tr. vestibulospinalis; 5 — tr. olivospinalis; 6 — tr. reticulospinalis; 7 — tr. spinocerebellaris anterior; S — tr. spinotlwlamicus lateralis; 9 — tr. rubrospinalis; 10 — tr. spinocerebellaris posterior; 11 — tr. corticospinalis lateralis; 12 — fasciculi proprii; 13 —fasciculus cuneatus; 14 — fasciculus gracilis; 15 — zona spongiosa; 16 — substantia gelatinosa; 17 — nuclei proprii. IS — columna thoracica; 19 — nucleus intermecliomedialis; 20 — nucleus interme- diolateralis; 21 — nuclei motorii
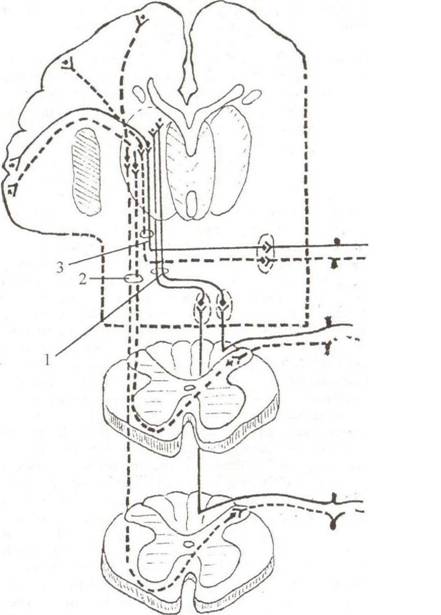
Проводящие пути сознательных проприоцептивных импульсов (tractus ganglio- bulbo-thalamo-corticalis). Пути Голля, Бурдаха(рис. 3.3)
Пути Голля и Бурдаха являются проводниками сознательного мышечно-суставного чувства от проприорецепторов аппарата движения. Первые нейроны представлены псевдоуниполярными клетками, тела которых лежат в спинномозговых узлах. Дендри- ты клеток направляются на периферию, где заканчиваются рецепторами в мышцах, сухожилиях, связках и капсулах суставов, костях и надкостнице. Аксоны клеток в составе задних корешков
Рис. 3.2. Проекционные проводящие пути внутренней капсулы: 1 — tr. frontothalamicus; 2 — tr.frontopontinus; 3 — tr. corticonuclearis; 4 — tr. corticospi- nalis, 5 — tr. thalamocorticalis; 6 — tr. occipitotemporopontinus; 7 — radiotio optica; 8 — radiatio acustica
вступают посегментно в спиннои мозг и, не входя в серое вещество, идут в восходящем направлении в составе задних канатиков, образуя тонкий пучок Голд я, fasciculus gracilis, и клиновидный пучок Бурдаха, fasciculus cuneatus. Путь Голля занимает внутреннее, а путь Бурдаха — наружное положение (см. рис. 3.1). Пучок Голля проводит глубокое мышечносуставное чувство от нижних конечностей и нижней половины туловища соответствующей стороны, включая волокна 19 нижних спинномозговых узлов, а пучок Бурдаха — от верхней части туловища, шеи и верхних конечностей, включая волокна 12 верхних спинномозговых узлов. Пучки Голля и Бурдаха достигают одноименных ядер (тонкого и клиновидного, nucl. gracilis и nucl. cuneatus), расположенных в дорсальных отделах продолговатого мозга, и здесь
Рис. з.з. Чувствительные проводящие пути коркового направления — пути Гэлля и Бурдаха; спинно-таламический путь: 1 — lemniscus medialis; 2 — lemniscus spinalis; 3 — lemniscus trigeminalis переключаются на вторые нейроны. В целом первые нейроны составляют путь — tractus gangliobulbaris. Аксоны вторых нейронов объединяются в единый пучок — tractus bulbothalamicus. Сначала нервные волокна проходят в вентромедиальном направлении по дугообразной траектории (они получили название внутренних дугообразных волокон, ft brae ar- cuatae internae). Затем волокна переходят на противоположную сторону и формируют компактный пучок, который совершает крутой изгиб, что и дало основание назвать его медиальной петлей, lemniscus medialis. На срединной линии происходит перекрест медиальных петель правой и левой сторон. Этот перекрест обозначается как чувствительный перекрест петель, decussatio lemniscorum (sensorum). Он располагается дорсальнее пирамид, между оливами, и формирует межоливный слой. Затем через дорсальную часть моста и покрышку ножек мозга волокна достигают таламуса, где переключаются на третьи нейроны в заднелатеральном вентральном ядре, nucl. ventralisposterolaterolis. В мосту к медиальной петле присоединяются пути кожной чувствительности шеи, туловища и конечностей (спинномозговая петля) и тройничная петля, образованная волокнами тройничного нерва, которые являются проводниками кожной и проприоцептивной чувствительности от области головы. Часть волокон вторых нейронов от клеток тонкого и клиновидного ядер направляется через нижнюю мозжечковую ножку к коре полушарий мозжечка одноименной стороны, другая часть — к коре полушарий мозжечка противоположной стороны. Благодаря этим связям мозжечок участвует в механизме координации движений. Аксоны третьих нейронов в составе tractus thalamocorticalis направляются к постцентральной извилине полушария большого мозга, где заканчиваются синапсами на клетках коры (IV слой). Волокна проходят через средний отдел задней ножки внутренней капсулы (см. рис. 3.2), а затем, веерообразно рассыпаясь, продолжают свой путь в составе лучистого венца. От нижней конечности и одноименной половины туловища сознательные проприоцептивные импульсы поступают в верхнюю треть постцентральной извилины, от верхней конечности — в среднюю, от головы — в нижнюю. Правой половине тела соответствуют извилины левого полушария большого мозга, а левой — правого. При выпадении глубокой проприоцептивной чувствительности нарушается представление о положении своего тела и его частей в пространстве, восприятие позы, ощущение активных и пассивных движений. Нарушается координация движений, утрачивается их ловкость и согласованность.
Проводящие пути кожного анализатора, tractus spinothalamicus lateralis, tractus spinothalamicus ventralis
Проводящие пути кожного анализатора состоят из проводящих путей болевой и температурной чувствительности, тактильной чувствительности и пространственной кожной чувствительности. Проводящий путь болевой и температурной чувствительности, tr. spinothalamicus lateralis (tractus ganglio-spirw- thalamo-corti- calis), проводит импульсы от экстерорецепторов кожи конечностей, туловища, шеи (рис. 3.3). Нервные импульсы болевой и температурной чувствительности от кожи лица проходят по нервным волокнам тройничного нерва. Первыми нейронами являются рецепторные псевдоуниполярные клетки, тела которых располагаются в спинномозговых узлах. Их периферические отростки заканчиваются экстерорецепторами в коже. Свободные нервные окончания воспринимают боль, тельца Руффини — тепло, колбочки Краузе — холод. Аксоны первых нейронов в составе задних корешков вступают в спинной мозг и делятся каждый Т-образно на восходящую и нисходящую ветви, от которых отходят множество коллатералей. Концевые разветвления и коллатерали направляются к заднему рогу и заканчиваются в нем синапсами на телах и дендри- тах клеток собственных ядер, nuclei proprii, которые являются вторыми нейронами. Совокупность аксонов первых нейронов образует tractus gangliospiitalis. Аксоны вторых нейронов совершают перекрест, постепенно переходя на протяжении всего спинного мозга на противоположную сторону через переднюю белую спайку, commissura alba. При этом они идут не горизонтально, а косо поднимаются на 2—3 сегмента выше уровня расположения тел нервных клеток, посылающих эти волокна, что необходимо учитывать при определении очага поражения. Далее, группируясь, аксоны вторых нейронов формируют латеральный спинно-таламический путь, tractus spinotlialamicus lateralis, который усиливается за счет Бологим нышерасположенных сегментов, и направляется в составе бокового канатика спинного мозга в головной (см. рис. 3.1). На уровне спинного мозга латеральный спинно-таламический пучок подразделяется на переднюю часть, проводящую болевую чувствительность, и заднюю, волокна которой проводят температурную чувствительность. В области продолговатою мозга латеральный спинно-таламический пучок располагается дорсаль- нее ядра оливы. Его волокна образуют спинномозговую петлю, lemniscus spinalis, которая сближается с медиальной петлей и следует с ней к зрительному бугру, где волокна переключаются на третьи нейроны в заднелатеральном вентральном ядре, nucl. ven- tralis posterolateralis. Аксоны третьих нейронов вместе с другими афферентными волокнами, проводящими общую чувствительность, проходят через среднюю часть задней ножки внутренней капсулы (см. рис. 3.2) и в составе таламо-кортикального пучка, tractus thala- mocorticalis, направляются к постцентральной извилине полушарий большого мозга (корковый конец кожного анализатора), где заканчиваются на клетках IV слоя коры. Проводящий путь тактильной чувствительности, tractus spi- nothalamicus ventralis, обеспечивает чувство осязания, прикосновения, давления. Первые нейроны представлены псевдоунипо- лярными клетками и лежат в спинномозговых узлах. Их дендри- ты заканчиваются экстерорецепторами в коже: осязательными тельцами (чувствительные тельца Мейснера), осязательными менисками (диски Меркеля), пластинчатыми тельцами (тельца Фатера-Пачини), которые воспринимают давление. Аксоны клеток спинномозгового узла в составе задних корешков вступают в спинной мозг и делятся каждый Т-образно на восходящую и нисходящую ветви, от которых отходят множество коллатералей, заканчивающихся в верхушке заднего рога на клетках студенистого вещества, substantia gelatinosa, которые являются телами вторых нейронов. Меньшая часть волокон рецепторных клеток, проводящих тактильную чувствительность, войдя в спинной мозг в составе заднего корешка, не заходит в серое вещество, а в составе задних канатиков поднимается до клеток тонкого и клиновидного ядер, следуя далее в составе медиальной петли. Аксоны вторых нейронов, тела которых располагаются в студенистом веществе, образуют перекрест, проходя через белую спайку спинного мозга на противоположную сторону. Затем в составе передних канатиков они направляются в головной мозг, образуя передний спинно-таламический тракт (см. рис. 3.1). Этот путь проходит через продолговатый мозг, затем — через дорсальную часть моста вместе с волокнами медиальной петли, через покрышку среднего мозга и заканчивается на телах третьих нейронов нис/. ventralis posterolateralis. Аксоны третьих нейронов проходят в составе tractus thalamo- corticalis через заднюю ножку внутренней капсулы и заканчиваются в постцентральной извилине и верхней теменной дольке, которые являются корковым концом кожного анализатора.
ПРОПРИОЦЕПТИВНЫЕ ПУТИ К МОЗЖЕЧКУ
Проприоцептивные пути к мозжечку составляют часть рефлекторного аппарата, осуществляющего функцию равновесия (бессознательную координацию движений). Они проводят бессознательное мышечно-суставное чувство от рецепторов аппарата движения, участвуют в регуляции мышечного тонуса.
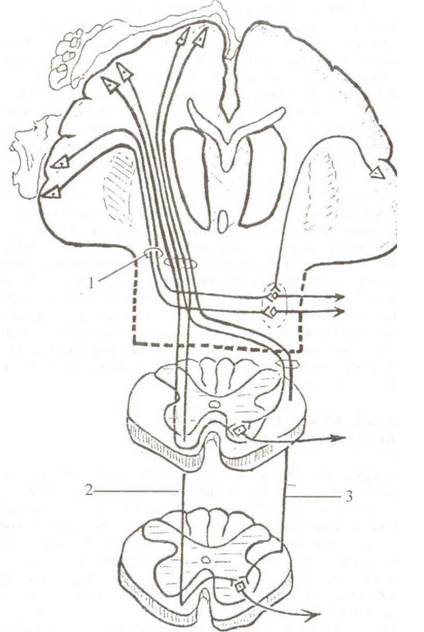
Передний спинно-мозжечковый путь, tractus spinocerebellaris ventralis (anterior), путь Говерса (рис. 3.4)
Тела первых нейронов располагаются в спинномозговых узлах. Их периферические отростки заканчиваются проприорецепто- рами в надкостнице, связках, капсулах суставов, мышцах. Центральные отростки вступают в спинной мозг в составе задних корешков и идут к промежуточной зоне серого вещества, заканчиваясь на телах и дендритах нейронов медиального промежуточного ядра, nucleus intermediomedialis (на вторых нейронах). Совокупность аксонов вторых нейронов образует передний спинно-мозжечковый путь. Большая часть волокон этого пути совершает перекрест, переходя через белую спайку на противоположную сторону, и в составе бокового канатика устремляется
Рис. 3.4. Проводящие пути проприоцептивной чувствительности к мозжечку: 1 — tractus spinocerebellaris dorsalis: 2 — tractus spinocerebellaris ventral is; 3 — nucleus ruber
в восходящем направлении (см. рис. 3.1). Оставшаяся часть волокон не совершает перекреста и проходит в составе боковых канатиков своей стороны. По мере восхождения путь усиливается за счет волокон из вышележащих сегментов. В продолговатом мозге он располагается между нижним оливным ядром и нижней мозжечковой ножкой. Далее передний спинно-мозжечковый путь проходит в дорсальном отделе моста и на границе со средним мозгом круто поворачивает в область верхнего мозгового паруса, где часть волокон переходит на противоположную сторону. Через верхние мозжечковые ножки волокна пути достигают коры червя мозжечка. В связи с тем, что нервные волокна дважды совершают перекрест (в белой спайке спинного мозга и в верхнем мозговом парусе), импульсы бессознательной проприоцептивной чувствительности передаются коре червя мозжечка преимущественно с одноименной стороны тела. Задний спинно-мозжечковый путь, tractus spinocerebellaris dorsalis (posterior), путь Флексига (рис. 3.4)
Тела первых нейронов располагаются в спинномозговых ганглиях. Периферические отростки клеток ганглиев заканчиваются проприорецепторами в мышцах, сухожилиях, капсулах суставов, связках. Центральные отростки в составе задних корешков вступают в спинной мозг и заканчиваются на нейронах грудного ядра, nucleus thoracicus (colunma f/ioracica) (вторых нейронах). Совокупность аксонов вторых нейронов составляет задний спинно-мозжечковый путь. Волокна пути устремляются в восходящем направлении, занимая периферическую часть заднего отдела бокового канатика спинного мозга (см. рис. 3.1). В продолговатом мозге волокна располагаются между нижним оливным ядром и волокнами спинномозгового пути тройничного нерва. Через нижние ножки мозжечка они следуют к коре червя. В связи с тем, что волокна заднего спинно-мозжечкового пути в продолговатом и спинном мозге не совершают перекреста, его называют прямым или неперекрещенным. Проприоцептивные пути мозжечкового направления и идущие к коре полушарий большого мозга дают возможность человеку тонко чувствовать свое тело и точно выполнять высокодифференцированные движения.
ДВИГАТЕЛЬНЫЕ ПРОВОДЯЩИЕ ПУТИ
Эфферентные пути проводят нервный импульс из различных центров головного мозга к рабочему органу (мышца, железа). У человека прямые эфферентные пути начинаются лишь из коры больших полушарий, где располагаются тела так называемых центральных нейронов. Эти пути являются проводниками сознательных двигательных импульсов на скелетную мускулатуру и объединяются понятием пирамидная система. Промежуточные ядра ствола (красное ядро, ядра крыши среднего мозга, ядра ретикулярной формации, ядро оливы, латеральное преддверное ядро) рассматриваются как начало эфферентных проводящих путей, которые посылают бессознательные двигательные им пульсы на скелетную мускулатуру и входят в состав экстранира- мидной системы. Мозжечок осуществляет связь со спинным мозгом через указанные стволовые ядра. Все нисходящие пути заканчиваются на телах и дендритах нейронов двигательных ядер передних рогов серого вещества спинного мозга или двигательных ядер черепных нервов (периферические нейроны). Таким образом, все эфферентные проводящие пути подразделяются на две группы: 1. Пирамидные пути. 2. Экстрапирамидные пути.
ПИРАМИДНЫЕ ПУТИ
Пирамидные пути проводят сознательные (волевые) двигательные импульсы, а также тормозные импульсы от коры полушарий большого мозга к нейронам двигательных ядер черепных нервов и к нейронам двигательных ядер передних рогов серого вещества спинного мозга. В зависимости от места назначения пирамидные пути подразделяются на две группы: • корково-ядерный путь, troctus corticonuclearis • корково-спинномозговой путь, tractus corticospinalis. Оба пути начинаются от пирамидных клеток предцентраль- ной извилины (V слой), нейронов околоцентральной дольки, задних отделов верхней и средней лобных извилин. Указанные области образуют так называемую двигательную зону коры. В преднентралыюй извилине проекция для мышц представлена в порядке, обратном их расположению в организме: верхним и парацентральным отделам соответствует нижняя конечность, средним — верхняя, нижним — область шеи и головы (рис. 3.5) — схема Пенфилда, «моторный гомункулус».
Корково -ядерный путь, tractus corticonuclearis (см. рис. 3.5)
Корково-ядерный путь образован аксонами нейронов, тела которых в виде крупных пирамидных клеток располагаются в нижнелатеральной трети предцентральной извилины. Аксоны
Рис. 3.5. Пирамидные пути: 1 — tr. corticonuclearis; 2 — tr. corticospinalis anterior;. 3 — tr. corticospincilis lateralis веерообразно сходятся в пучок, который занимает область колена внутренней капсулы (см. рис. 3.2). Пройдя промежуточный мозг снаружи от зрительного бугра, корково-ядерный путь направляется в основание ножки мозга той же самой стороны и располагается в ее средней части. В среднем мозге часть волокон отделяется от основного пучка и направляется в покрышку ножек мозга, заканчиваясь на клетках ядер глазодвигательного (III) и блокового (IV) нервов как своей, так п противоположной стороны. Большая часть волокон изоснования ножки переходит в основание моста. Здесь от корково-ядерного пути вновь отделяются волокна, которые заканчиваются на нейронах двигательных ядер тройничного (V), отводящего (VI) и лицевого (VII) нервов. Оставшаяся часть волокон, достигая продолговатого мозга, заканчивается на мотонейронах двигательных ядер языкоглоточного (IX), блуждающего (X), добавочного (XI) и подъязычного (XII) нервов. Большая часть ядер получает волокна от обоих полушарий, за исключением нижней части ядра VII пары и ядра XII пары, где заканчиваются нервные волокна только из противоположного полушария. Нейроны двигательных ядер черепных нервов являются эффекторными клетками корковоядерного пути, проводящими двигательные импульсы к поперечнополосатым мышцам головы, некоторым мышцам шеи, мышцам глотки и гортани.
Корково -спинномозговой путь, tractus corticospinalis (см. рис. 3.5)
Эфферентные проекционные волокна, входящие в состав пирамидной системы, берут начало от пирамидных нейронов пятого слоя коры больших полушарий. При этом около 40% этих волокон начинаются от гигантских пирамидных клеток коры предцентральной извилины и парацентральной дольки, 20% — от пирамидных нейронов коры постцентральной извилины и 40% — от пирамидных нейроцитов задней трети верхней и средней лобных извилин, передней трети верхней теменной дольки и надкраевой извилины головного мозга. Аксоны первых нейронов входят в состав лучистого венца и идут в нисходящем направлении, занимая во внутренней капсуле передние две трети задней ножки (см. рис. 3.2). Пройдя про межуточный мозг сбоку от зрительных бугров, волокна корково-спинномозгового пути занимают средний отдел основании ножек мозга снаружи от tractus corticonuclearis. В заднем мозге корково-спинномозговой путь рассыпается на множество пучков, которые находятся в основании моста, и отделяются друг от друга многочисленными собственными ядрами моста. В продолговатом мозге волокна пирамидного пути вновь сближаются и образуют пирамиды, расположенные на его вентральной поверхности. На границе со спинным мозгом большая часть волокон каждой пирамиды (80%) переходит на противоположную сторону, образуя пирамидный перекрест, decussatio pyramidum. Этот перекрест также получил название двигательного, в отличие от верхнего, чувствительного, перекреста петель. Волокна, перешедшие на противоположную сторону, направляются в боковой канатик спинного мозга. Остальные волокна идут в передний канатик спинного мозга своей стороны (см. рис. 3.1). Таким образом, происходит разделение единого до сих пор корковоспинномозгового пути на два самостоятельных в спинном мозге тракта: tractus corticospinalis lateralis и tractus corticospinalis anterior. Волокна латерального пирамидного пути заканчиваются на клетках двигательных ядер передних рогов серого вещества каждого сегмента спинного мозга до его крестцового отдела. Причем каждое волокно заканчивается на нейронах сразу нескольких сегментов. Наиболее значительное количество волокон отделяется от бокового пирамидного пути в области утолщений спинного мозга, что соответствует большему количеству эффектор- ных нейронов в этих сегментах, обеспечивающих иннервацию многочисленных мышц верхних и нижних конечностей. Волокна переднего корково-спинномозгового пути в области передней белой спайки посегментпо переходят на противоположную сторону, заканчиваясь на нейронах двигательных ядер передних рогов. Постепенно истончаясь, передний корково-спинномозговой путь заканчивается на уровне среднего грудного отдела спинного мозга. Волокна корково-спинномозгового пути по ходу отдают множество коллатералей. Так, часть коллатералей заканчивается на нейронах собственных ядер моста, аксоны которых образуют мосто-мозжечковый путь. Его волокна преимущественно переходят на противоположную сторону. Благодаря этим связям осуществляется мозжечковая поправка на все виды произвольных движений. Другая часть волокон пирамидных путей достигает зрительного бугра, базальных ядер, красного ядра, черной субстанции. Аксоны вторых нейронов покидают спинной мозг в составе передних корешков и по спинномозговым нервам направляются к скелетной мускулатуре шеи, туловища и конечностей. Иннервация мускулатуры является перекрестной. Правое полушарие обеспечивает произвольные движения левой половины тела и наоборот. От сложности движений, выполняемых различными группами мышц, зависит величина соответствующей области коркового центра. Наиболее крупные по размерам участки коры связаны с мышцами, которые обеспечивают особо сложные и дифференцированные движения. Так, мышцам кисти и пальцев соответствует территория, занимающая почти всю среднюю треть предцентральной извилины. Полное поражение пирамидных путей в пределах головного или спинного мозга ведет к развитию центрального паралича. При нарушении целостности аксона или тела двигательною нейрона спинного мозга возникает периферический паралич. Для центрального паралича характерны мышечная гипертония, повышение сухожильных рефлексов и появление патологических рефлексов. Парализованные мышцы не подвергаются дегенеративной атрофии. Поражение периферического нейрона вызывает атонию, арефлексию и атрофию.
ПРОВОДЯЩИЕ ПУТИ ЭКСТРАПИРАМИДНОИ СИСТЕМЫ
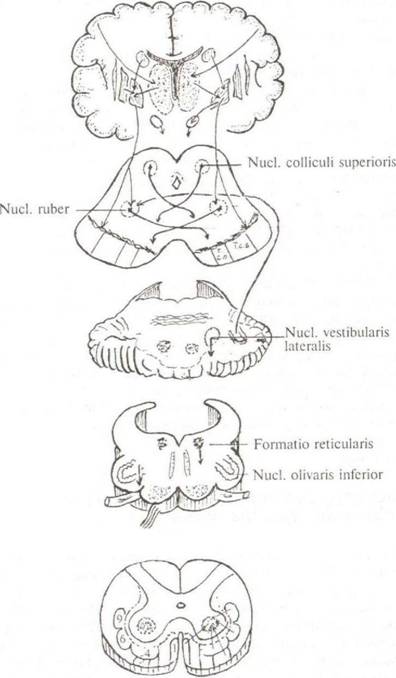
Экстрапирамиднии система — это система подкорковых п стволовых ядер головного мозга и двигательных внепирамидных проводящих путей, осуществляющая непроизвольную автоматическую регуляцию и координацию сложных двигательных актов, регуляцию мышечного тонуса, поддержание позы, организацию двигательных проявлений эмоций (рис. 3.6). К экстрапирамидной системе относятся базальные ядра (хвостатое, чечевицеобразное, ограда), медиальные ядра таламуса, субталамическое ядро (люисово тело), ядра крыши среднего мозга, черное вещество, красное ядро, ядра ретикулярной формации, вестибулярные ядра, нижнее оливное ядро, мозжечок и
Рис. 3.6. Проводящие пути экстра пирамидной системы волокна белого вещества, осуществляющие многочисленные связи между этими ядрами и спинным мозгом. Среди структур экстрапирамидной системы центральное положение занимает так называемая «стриопадлидарная система». При этом хвостатое ядро и скорлупу чечевицеобразного ядра объединяют под названием «стриатум», а бледный шар обозначают как «паллмдум». К паллидарной системе относят также черное вещество и красное ядро. Стриопадлидарная система отвечает за регуляцию таких сложных двигательных актов, как ходьба, бег, плавание и др., которые осуществляются с участием различных мышечных групп. При этом «стриатум» ведает организацией движений, обеспечивая их автоматизм, энергетическую расчетливость, скупость, в то время как «паллидум», координируя тонус и фазовую двигательную активность мышц, обеспечивает движения мощные, точные, но энергетически расточительные. Стриопадлидарная система должна получать исчерпывающую информацию о состоянии мышц, сухожилий, суставов, о положении тела в пространстве и т. д. Афферентные связи стриопалли- дарной системы включают таламус, мозжечок, ретикулярную формацию, кору больших полушарий. Эфферентные волокна направляются к стволовым ядрам экстрапирамидной системы. От стволовых ядер начинаются пути к двигательным нейронам передних рогов серого вещества спинного мозга: красно- ядерно-спинномозговой; покрышечноспинномозговой; ретику- ло-спинномозговой; преддверноспинномозговой; оливо-спинномозговой. Красноядерно-спинномозговой путь, tractus rubrospinalis, монаков путь Начинается от клеток красного ядра, расположенного в покрышке среднего мозга. Аксоны мультиполярных нейронов, покинув ядро, тотчас переходят на противоположную сторону, образуя вентральный перекрест покрышки, decussatio tegmentalis ventralis (перекрест Фореля). Далее волокна проходят через ретикулярную формацию моста и продолговатого мозга, спускаются в боковые канатики спинного, располагаясь кпереди от латерального корково-спинномозгового пути (см. рис. 3.1), и посегментно заканчиваются на мотонейронах двигательных ядер пе- редких рогов серого вещества спинного мозга. Красное ядро считается основным координационным центром экстрапирамидной системы. Ог мозжечка через красноядерно- спинномоз- говой путь следуют импульсы, осуществляющие так называемую мозжечковую поправку, которая предотвращает проявление инерции движущегося тела. Покрышечно -спинномозговой путь, tractus tectospinalis Тела первых нейронов располагаются в глубоких слоях верхних бугорков крыши среднего мозга. Их аксоны огибают центральное серое вещество водопровода мозга и переходят на противоположную сторону, образуя дорсальный перекрест покрышки или фонтановидный перекрест, decusscitio tegmentalis dorsalis (перекрест Меймерта). Затем идут в нисходящем направлении, располагаясь в дорсальной части моста и продолговатого мозга. Опускаясь в спинной мозг, проходят в составе передних канатиков, где располагаются вплотную к передней срединной щели (см. рис. 3.1). Аксоны первых нейронов заканчиваются на телах двигательных нейронов шейных сегментов спинного мозга. Часть нервных волокон этого пути заканчивается на ядрах глазодвигательных нервов и выделяется под названием «покрышечно-бульбарные волокна». Покрышечноспинномозговой путь осуществляет бессознательные двигательные реакции в ответ на слуховые и зрительные раздражения, обеспечивая защитный рефлекс на- стораживания, который выражается в повороте головы в сторону светового или звукового раздражителя. Причем реакция на акустические стимулы обусловлена связями верхних и нижних бугорков четверохолмия.
Ретикуло-спинномозговой путь, tractus reticulospinalis
Первые нейроны ретикуло-спинномозгового пути располагаются в ретикулярной формации мозгового ствола. Аксоны муль- типолярных нейронов идут в нисходящем направлении и, собираясь в пучок, проходят в составе передних канатиков спинного мозга, занимая его наружные отделы (см. рис. 3.1), и посегмент- но заканчиваются на двигательных нейронах передних рогов. Ретикуло-спинномозговой путь служит для выполнения сложных рефлекторных реакций организма, требующих для их осуществления одновременного участия многих групп поперечно-полосатых мышц. Он оказывает активирующее влияние, которое клинически проявляется повышением мышечного тонуса, и тормозящее, приводящее к снижению мышечного тонуса.
Преддверно -спинномозговой путь, tractus vestibulospinalis Путь начинается в дорсальной части моста из латерального (Дейтерса) и нижнего (Роллера) вестибулярных ядер. В виде небольшого пучка он проходит позади олив продолговатого мозга, а затем на границе переднего и бокового канатиков спинного мозга (см. рис. 3.1) и заканчивается на нейронах двигательных ядер шейного, грудного и поясничного отделов спинного мозга. Путь связывает мозжечок и вестибулярные ядра с мышцами обеих половин тела, осуществляя проведение импульсов, направленных на сохранение равновесия тела и регуляцию мышечного тонуса.
Оливо-спинномозговой путь, tractus olivospinalis, Бехтерева-Гельвега Путь начинается от нижнего оливного ядра продолговатого мозга. Аксоны мультпполярных нейронов собираются в пучок, который составляет передненаружный отдел боковых канатиков спинного мозга (см. рис. 3.1). Посегментно он заканчивается на нейронах двигательных ядер передних рогов четырех верхних шейных сегментов спинного мозга. Путь обеспечивает безусловнорефлекторное поддержание тонуса мышц шеи и двигательные акты, направленные на сохранение равновесия тела.
|
||
|
|
Последнее изменение этой страницы: 2018-05-10; просмотров: 886. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |