|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
ФУНКЦИОНАЛЬНАЯ АНАТОМИЯ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
КРАТКИЕ СВЕДЕНИЯ О СТРУКТУРНОЙ ОРГАНИЗАЦИИ НЕРВНОЙ ТКАНИ
Основные структурные компоненты нервной ткани — это нервные клетки (нейроны) и нейроглия. Нейроны определяют главные свойства нервной ткани — возбудимость и проводимость. Нейроглия обеспечивает условия существования и функциональной активности нейронов, т. е. выполняет опорную, защитную, трофическую и секреторную функции. Нейроны. Общее количество нейронов в нервной системе человека составляет около 10м, причем каждая клетка способна устанавливать до 10 000 контактов с другими нейронами, что определяет разнообразие и сложность нервной деятельности. Как любая клетка, нейрон имеет клеточное тело (перикарион), а его специфической особенностью является наличие специали- шрованных отростков (рис. 2.1).
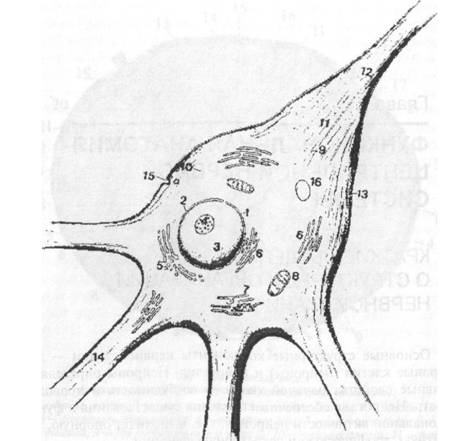
Клеточные тела нейронов имеют самую разнообразную форму. Различают пирамидные, звездчатые, веретенообразные, кор- шпчатые, горизонтальные и даже паукообразные нейроны. Размеры нервных клеток колеблются от 4—6 до 140 мкм. Клеточное тело нейрона состоит из клеточной оболочки, ядра и цитоплазмы. Клеточная оболочка(нейролемма) отделяет клетку от окру-
Рис. 2.1. Улыпраструктурпая организация нейрона (схематично): 1 — ядерная оболочка; 2 — ядерные поры; 3 — кариоплазма; 4 — ядрышко; 5 — аппарат Гольджи; 6 — агранулярная эндоплазматическая сеть; 7 — гранулярная эндоплазматическая сеть; 8 — митохондрия; 9 — микротрубочки; 10 — микрофиламен- ты; 11 — аксонный холмик; 12 — начальная часть аксона; 13 — перикарион; 14 — дендриты; 15 — пиноиитозные пузырьки; 16 — липофусциновые гранулы
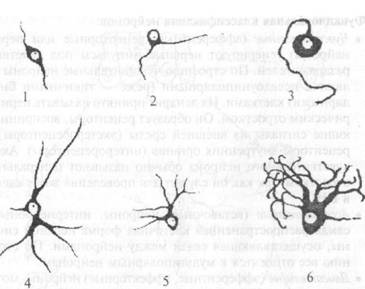
жающей среды и выполняет рецепторную и транспортную функции. Нейролемма обладает способностью к проведению нервного импульса (распространению деполяризации). Ядро представляет собой систему генетической детерминации и регуляции белкового синтеза. Цитоплазма (нейроплазма) состоит из гиалоплазмы (внутренняя среда клетки) с расположенными в ней органеллами и включениями.  Органеллы являются обязательными для всех клеток микроструктурами и выполняют специализированные жизненно важные функции. Они подразделяются на мембранные и немембранные. К мембранным органеллам относятся эндоплазматическая сеть, аппарат Гольджи, лизосомы и митохондрии. Эндоплазматическая сеть участвует в процессах синтеза и внутриклеточного гранспорта веществ. По строению различают гранулярную и агранулярную эндоплазматическую сеть. Гранулярной называется шдоплазматическая сеть с фиксированными рибосомами. Она принимает участие в синтезе белка. В нервных клетках гранулярная эндоплазматическая сеть хорошо развита. Ее уплощенные цистерны образуют особые комплексы, которые на светооптическом уровне имеют вид базофильных глыбок или зерен различных размеров и формы. Они получили название хроматофильного вещества (субстанции Ниссля) или тигроида. Характер распределения и размеры шыбок хроматофильного вещества являются показателем активности белково-синтетических процессов в клетке. На мембранах агранулярной сети рибосомы отсутствуют. Ее функция связана с метаболизмом липидов и некоторых полисахаридов. Аппарат Гольджи хорошо развит в нервных клетках. Он обеспечивает накопление продуктов, синтезированных в эндоплазматической сети, их созревание, преобразование и выведение готовых секретов за пределы клетки. Лизосомы определяют процессы внутриклеточного переваривания, т. е. расщепления различных биопо- чимеров. Лизосомальный аппарат нервных клеток характеризуется высокой активностью. Митохондрии являются органеллами клеточного дыхания. Их функция связана с окислением органических соединений и использованием освобождающейся при распаде этих соединений энергии для синтеза молекул АТФ. Н нервных клетках митохондрии очень многочисленны и обеспечивают высокие энергетические потребности нейрона. К немембранным органеллам относятся рибосомы и полисомы, а также фибриллярные компоненты цитоплазмы. Рибосомы и полисомы обеспечивают белково-синтетические процессы. Фибриллярными компонентами цитоплазмы являются микротрубочки, микрофиламенты и микрофибриллы (промежуточные фила- менты). Они являются опорным аппаратом клетки, образуют ее иптоскелет. Цитоскелет нейрона хорошо развит и представлен всеми элементами, образующими трехмерную опорно-сократи тельную сеть, которая способствует сохранению формы клетки и аксона. На светооптическом уровне элементы цитоскелета нервной клетки выявляются в виде так называемых нейрофибрилл при окрашивании солями серебра. Нейрофибриллы образованы промежуточными филаменгами и микротрубочками, которые склеиваются в пучки при фиксации. В цитоплазме они образуют плотную сеть, а в волокнах ориентированы параллельно. Включения являются необязательным компонентом клетки. Они появляются и исчезают в зависимости от ее функционального состояния. В цитоплазме нейрона включения представлены липидными каплями, гранулами липофусцина, меланина. Отростки нейронов представляют собой тонкие цитоплазматические выросты в виде нитей. По функциональному назначению они делятся на два вида: аксоны и дендриты. Аксон (от греч. ахоп — ось), или нейрит, выполняет функцию отведения нервного импульса от тела клетки. Аксон заканчивается концевым аппаратом на другом нейроне или на клетках рабочего органа. Любая нервная клетка имеет только один аксон. Аксон может содержать до 99% объема цитоплазмы. Участок тела клетки, от которого отходит аксон, называется аксонным холмиком, в котором генерируются нервные импульсы. Аксоны могут отдавать ветви (аксональные коллатерали). Они отходят почти под прямым углом, а затем поворачивают на 90° и продолжают идти вдоль аксона в прежнем или обратном (возвратная коллатераль) направлении. Дендрит (от греч. dendron — дерево) проводит нервный импульс по направлению к телу клетки. В большинстве случаев эти отростки сильно ветвятся. Количество, длина и характер ветвления дендритов специфичны для разных типов нейронов. Обычно дендриты несколько раз делятся дихотомически. В функциональной нейроморфологии широко применяются две классификации нейронов: 1. Морфологическая классификация по числу отростков (рис. 2.2): • Униполярные нейроны — клетки с одним отростком. В постнатальном периоде онтогенеза человека подобные клетки не встречаются. В период внутриутробного развития в стенке нервной трубки зародыша обнаруживаются униполярные нейробласты.
Рис. 2.2. Типы нервных меток: 1 — биполярный нейрон: 2 — псевдоуниполярный нейрон; 3 — униполярный нейрон;4,5,6 — различные формы мультиполярных нейронов • Биполярные нейроны — клетки с двумя отростками. Аксон и дендрит обычно отходят от противоположных полюсов клетки. В нервной системе человека к типичным биполярным клеткам относятся нейроны сетчатки глаза, вестибулярного и спирального ганглиев. Разновидностью биполярных нейронов являются псевдоуниполярные нейроны. Аксон и дендрит таких нейронов отходят от клеточного тела в виде общего цитоплазматического выроста, создающего впечатление одного отростка, с последующим Т-образным его разделением. Псевдоуниполярные нейроны обнаруживаются в спинальных и краниальных чувствительных ганглиях. • Мультиполярные нейроны — клетки с тремя или более отростками, один из которых — аксон, остальные — дендриты. Такие нейроны — самая распространенная клеточная форма в нервной системе человека. По длине аксона мультипо- лярные нейроны делятся на две группы: клетки Гольджи I типа (имеют длинный аксон) и клетки Гольджи II типа (имеют короткий аксон). 2. Функциональная классификация нейронов: • Чувствительные (афферентные, рецепторные или первые нейроны) генерируют нервные импульсы под действием раздражителей. По строению чувствительные нейроны являются псевдоуниполярными (реже — типичными биполярными) клетками. Их дендрит принято называть периферическим отростком. Он образует рецепторы, воспринимающие сигналы из внешней среды (экстерорецепторы),. и рецепторы внутренних органов (интерорецепторы). Аксон чувствительного нейрона обычно называют центральным отростком, так как он служит для проведения возбуждения в ЦНС. • Ассоциативные (вставочные) нейроны, интернейроны — самая распространенная клеточная форма нервной системы, осуществляющая связи между нейронами. По строению все относятся к мультиполярным нейронам. • Двигательные (эфферентные, эффекторные) нейроны, мотонейроны передают нервный импульс на рабочий орган. Нейроглия. Нейроглия (от греч. neuron — нерв, glia — клей) — совокупность глиальных клеток (глиоцитов). Нейроглия подразделяется на макроглию и микроглию. Макроглия включает астро- цитарную глию (астроциты, астроглпя), олигодендроглию (оли- годендрониты) и эпендимную глию (эпендимоциты), которые являются производными нейрального зачатка (рис. 2.3). Эпендимоциты выстилают полости желудочков головного мозга и центральный канал спинного мозга. В области сосудистых сплетений желудочков мозга они участвуют в образовании гема- толикворного барьера, через который происходит ультрафильтрация крови с образованием спинномозговой жидкости. Астроциты (от греч. astron — звезда, cytos — клетка) встречаются во всех отделах нервной системы. Астроциты подразделяются на две группы: протоплазматические и волокнистые. Про- топлазматические астроциты встречаются преимущественно в сером веществе ЦНС. Волокнистые астроциты располагаются главным образом в белом веществе. Основные функции астроглии — опорная, метаболическая, барьерная и защитная. Эти клетки образуют опорный каркас ЦНС, внутри которого располагаются другие клетки и волокна. Метаболическая функция направлена на поддержание ионного и медиаторного баланса.
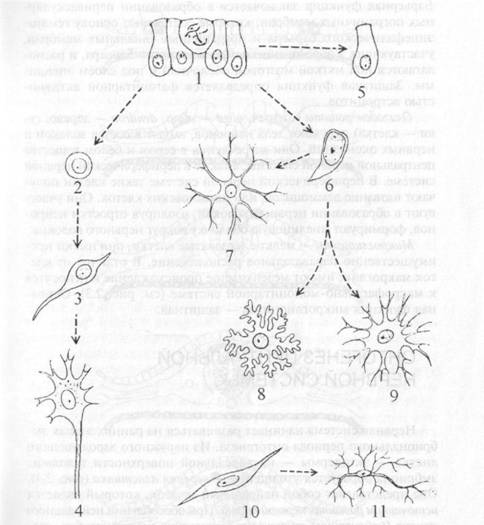
Рис. 2.3.Гистогенез нейронов и нейроглии; типы глиальных клеток: I — нейроэпителиальные клетки; 2 — аполярный нейробласт; 3 — биполярный ней- робласт; 4 — мультиполярный нейробласт; 5 — эпендимоцит; 6— глиобласт; 7— олигодендроцит; 8—протиплазматический астроцит; 9 — волокнистый остропит; 10 — клетка мезенхимы; II — микроглия Барьерная функция заключается в образовании периваскуляр- ных пограничных мембран, которые составляют основу гематоэнцефалического барьера и пограничных глиальных мембран, участвующих в образовании нейроликворного барьера, и располагаются под мягкой мозговой оболочкой и под слоем эпендимы. Защитная функция определяется фагоцитарной активностью астроцитов. Олигодендроциты (от греч. oligo — мало, dendron — дерево, су- tos — клетка) окружают тела нейронов, входят в состав волокон и нервных окончаний. Они встречаются в сером и белом веществе центральной нервной системы, а также в периферической нервной системе. В периферической нервной системе такие клетки получают название леммоцитов, или шванновских клеток. Они участвуют в образовании нервных волокон, изолируя отростки нейронов, формируют миелиновую оболочку вокруг нервного волокна. Микроглиоциты — мелкие звездчатые клетки, они имеют преимущественно паравазальное расположение. В отличие от клеток макроглии, имеют мезенхимное происхождение и относятся к макрофагально-моноцитарной системе (см. рис. 2.3). Основная функция микроглиоцитов — защитная.
ОНТОГЕНЕЗ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
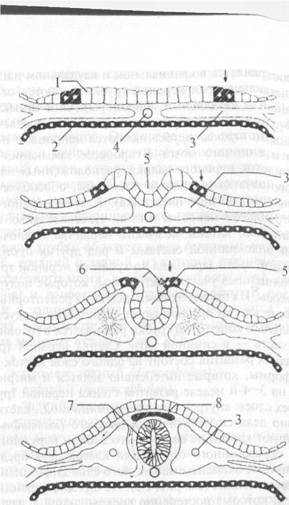
Нервная система начинает развиваться на ранних этапах эмбрионального периода онтогенеза. Из наружного зародышевого листка — эктодермы — на дорсальной поверхности туловища эмбриона образуется утолщение — нервная пластинка (рис. 2.4). Она представляет собой нейральный зачаток, который является источником развития нервной ткани. При обособлении нейрального зачатка (нейруляции) происходит образование нервной трубки, нервного гребня и нейральных плакод. На 18—21-е сутки развития эмбриона человека нервная пластинка превращается в нервный желобок. Приподнятые края нервного желобка называются нервными валиками. На 22е сутки нервный желобок замыкается в нервную трубку и обособляется от эктодермы. Этот процесс начинается в шейном отделе, а за-
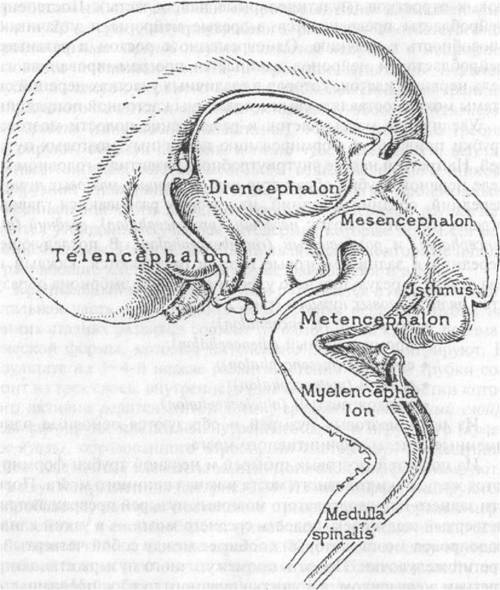
Рис. 2.4. Гистогенез нервной ткани: 1 — нейроэктодерма (нервная пластинка); 2 — энтодерма; 3 — мезодерма; 4 — хорда; 5 — нервный желобок; 6 — нервные валики; 7— нервная трубка; 8 — нервный гребень; 9 — спинальный ганглий тем распространяется в краниальном и каудальном направлениях. Первоначально края нервной трубки остаются открытыми (краниальный и каудальный нейропоры). Они замыкаются на 24-е и 26-е сутки внутриутробного развития соответственно. Производными нервной трубки являются нейроны и нейроглия головного и спинного мозга. В процессе замыкания нервной трубки из клеток нервного валика, расположенных на границе нейрального зачатка и кожной эктодермы, обособляется нервный гребень (ганглиозная пластинка). Клетки нервного гребня мигрируют, образуя нейроны и глию чувствительных и вегетативных ганглиев, клетки мозгового вещества надпочечников, диффузной эндокринной системы и ряд других производных. В краниальной части зародыша по краям от нервной трубки находятся утолщенные участки эктодермы, которые получили название плакоды. Их производными являются рецепторные и поддерживающие клетки органов слуха, равновесия и вкуса. Краниальный конец этой трубки развивается в головной мозг, остальная часть — в спинной мозг. Стенка нервной трубки на ранних стадиях развития состоит из одного слоя клеток призматической формы, которые интенсивно делятся и мигрируют. В результате на 3—4-й неделе развития стенка нервной трубки состоит из трех слоев: внутреннего (эпендимный слой), клетки которого активно делятся митотически; среднего (мантийный слой), куда мигрируют клетки из внутреннего слоя, и наружного (краевая вуаль), образованного отростками клеток двух предыдущих слоев. Клеточные элементы мантийного слоя дифференцируются в двух направлениях (см. рис. 2.3). Из них дифференцируются нейробласты, которые постепенно превращаются в зрелые нервные клетки, и спонгиобласты (глиобласты), дающие начало аст- роцитарной глии и олигодендроглии. К третьему месяцу внутриутробной жизни начинается миграция нейробластов к месту назначения. Нейробласты перемещаются вдоль отростков глиальных клеток (радиальная глия), которые протягиваются через всю стенку нервной трубки. В дальнейшем радиальные глиоциты превращаются в астроциты. Миграция нейробластов продолжается до 32-й недели внутриутробной жизни. В ходе миграции нейробласты дифференцируются в нейроны (см. рис. 2.3). Сначала нейробласты не имеют отростков (аполярные нейробласты). Затем происходит формирование отростков на противоположных концах клеточных тел, в результате образуются биполярные нейробла- сты. Один из отростков таких клеток подвергается обратному развитию, а на его месте впоследствии появляются несколько новых отростков (мультиполярные нейробласты). Постепенно нейробласты превращаются в зрелые нейроны и утрачивают способность к делению. Одновременно с ростом и развитием пейробластов и нейронов начинается программированная гибель нервных клеток, которая в различных участках нервной системы может составлять свыше половины клеточной популяции. Увеличение массы клеток и расширение полости мозговой грубки приводит к формированию первичных мозговых пузырей. На третьей неделе внутриутробного развития в головном отделе нервной трубки образуется три первичных мозговых пузыря: передний, средний и задний, из которых развиваются главные отделы головного мозга: передний (prosencephalon), средний (mesencephalon) и ромбовидный (rhombencephalon). В последующем передний и задний мозговые пузыри разделяются, каждый на два отдела, в результате чего у 4—5-недельного эмбриона образуется пять мозговых пузырей (рис. 2.5): ♦ конечный (telencephalon); ♦ промежуточный (diencephalon); ♦ средний (mesencephalon); ♦ задний (metencephalon); ♦ продолговатый (myelencephalon). Из пяти мозговых пузырей и образуются основные одноименные отделы дефинитивного мозга. Из полостей мозговых пузырей и нервной трубки формируются желудочки головного мозга и канал спинного мозга. Полости заднего и продолговатого мозговых пузырей превращаются в четвертый желудочек, полость среднего мозга — в узкий канал, водопровод мозга, который сообщает между собой четвертый и третий желудочки. Полость промежуточного пузыря становится третьим желудочком, а полость конечного пузыря превращается в два боковых желудочка. Через посредство парного межжелудочкового отверстия третий желудочек сообщается с каждым боковым. Четвертый желудочек сообщается с каналом спинного мозга. В желудочках и спинномозговом канале циркулирует спинномозговая жидкость. К третьему месяцу внутриутробного развития выделяются все основные части центральной нервной системы: большие полушария и ствол мозга, мозговые желудочки, спинной мозг. К пятому
Рис. 2.5. Онтогенез головного мозга: стадия пяти мозговых пузырей месяцу дифференцируются основные борозды коры больших полушарий. Скорость роста отдельных частей нервной трубки различна, что приводит к образованию нескольких изгибов в ее краниальном отделе, которые в последующем развитии эмбриона исчезают. Однако в области соединения среднего и промежуточного мозга остается изгиб мозгового ствола под углом 90°. Головной мозг новорожденного имеет относительно большую величину, и масса его в среднем составляет около 400 г (Vs массы тела). У новорожденного хорошо выражены крупные борозды и извилины, однако их глубина и высота невелики. Мелкие бороздки появляются постепенно в течение первых лет жизни. К девяти месяцам первоначальная масса мозга удваивается и составляет 1/i 1 —V12 массы тела, к трем годам — утраивается (1 /1 з массы тела). К 20 годам первоначальная масса мозга увеличивается в 4—5 раз и составляет около 'До массы тела. Рост мозга происходит в основном за счет миелинизации нервных проводников и увеличения размеров уже имеющихся при рождении примерно 20 млрд нервных клеток. После рождения претерпевает изменения и спинной мозг. По сравнению с головным мозгом спинной мозг новорожденно- ю является более завершенным в структурном и функциональном отношении. К моменту рождения спинной мозг относительно длиннее, чем у взрослого. В дальнейшем рост спинного мозга отстает от роста позвоночника, в связи с чем его нижняя iparnma смещается в проксимальном направлении. Окончательное соотношение спинного мозга и позвоночного канала устанавливается к пяти — шести годам. Рост спинного мозга продолжается приблизительно до 20 лет. Его масса за это время увеличивается в 8 раз.
Спинной мозг
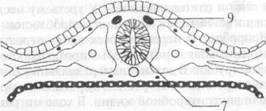
Внешнее строение. Спинной мозг, medulla spinalis, располагается в позвоночном канале. Он начинается на уровне I шейного позвонка и заканчивается на уровне II поясничного позвонка. Особенности внешнего строения характеризуются наличием следующих структурных образований (рис. 2.6):
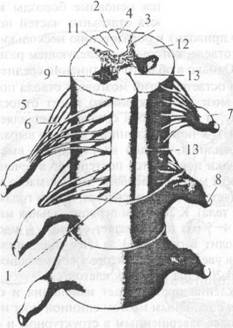
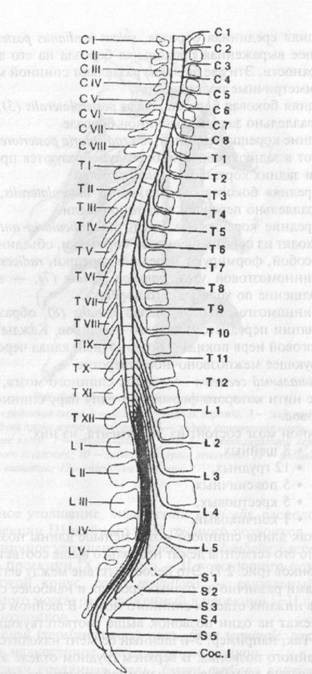
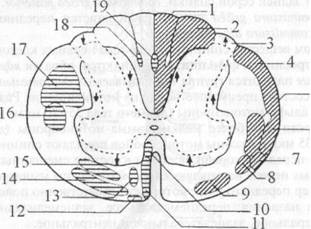
Рис. 2.6. Внешнее строение спинного мозга: 1 — передняя срединная щель; 2 — задняя срединная борозда; 3 — задняя боковая борозда; 4 — задняя промежуточная борозда; 5 — задние корешковые нити; 6 — передние корешковые нити; 7 — спинномозговой узел; 8 — спинномозговой нерв; 9 — передний рог серого вещества; 10 — задний рог серого вещества; 11 — задний канатик; 12 — боковой канатик; 13 — передний канатик • Шейное утолщение, intumescenlia cetyicalis, расположенное в проекции III шейного — II грудного позвонков. • Поясничное утолщение, intumescenlia lumbalis, расположенное в проекции IX грудного — II поясничного позвонков. • Мозговой конус, conus medullaris, — каудальный конец спинного мозга, расположенный в проекции I-II поясничных позвонков. • Концевая (терминальная) нить, filum terminale, заканчивается в надкостнице копчиковых позвонков. • Передняя срединная щель, fissura mediana anterior (1), — глубокая продольная щель на передней поверхности спинного мозга. • Задняя срединная борозда, sulcus medianus posterior (2), — менее выраженная продольная борозда на его задней поверхности. Эти две борозды разделяют спинной мозг на две симметричные половины. • Задняя боковая борозда, sulcus posterolateralis (3), проходит параллельно задней срединной борозде. • Задние корешковые нити, fila radiculariaposteriores (5), вступают в заднюю боковую борозду; образуются при разделении задних корешков, radices dorsales. • Передняя боковая борозда, sulcus anterolateralis, проходит параллельно передней срединной щели. • Передние корешковые нити, fila radicularia anteriores (6), выходят из передней боковой борозды и, объединяясь между собой, формируют передние корешки, radices anteriores. • Спинномозговой узел, ganglion spinale (7), — небольшое утолщение по ходу заднего корешка. • Спинномозговой нерв, nervus spinalis (8), образуется при слиянии переднего и заднего корешков. Каждый спинномозговой нерв покидает позвоночный канал через соответствующее межпозвоночное отверстие. • Спинальный сегмент — участок спинного мозга, корешковые нити которого формируют одну пару спинномозговых нервов. Спинной мозг состоит из 31 сегмента, из них: ♦ 8 шейных, ♦ 12 грудных, ♦ 5 поясничных, ♦ 5 крестцовых, ♦ 1 копчиковый. Так как длина спинного мозга меньше длины позвоночного канала, то его сегменты лежат несколько выше соответствующих им позвонков (рис. 2.7). Это несоответствие между сегментами и позвонками различно на разных уровнях и наиболее сильно выражено в нижних отделах спинного мозга. В шейном отделе сегменты лежат на один позвонок выше соответствующего им по номеру. Так, например, 5-й шейный сегмент находится на уровне IV шейного позвонка. В верхнем грудном отделе это несоответствие равно двум, т. е. тот или иной сегмент лежит на уровне не одноименного с ним по счету позвонка, а на два уровня выше. В нижнем грудном отделе оно достигает трех позвонков.
Рис. 2.7. Топография спинальных сегментов. В итоге грудной отдел спинного мозга оканчивается на уровне верхнего края X грудного позвонка. Поясничные сегменты лежат на уровне X, XI и частично XII грудных позвонков, крестцовые — на уровне XII грудного и верхнего края II поясничного позвонков. Ниже последнего в позвоночном канале находятся только корешки поясничных и крестцовых нервов. Они проходят почти параллельно продольной оси спинного мозга, так что мозговой конус и терминальная нить оказываются расположенными среди густого пучка нервных корешков, который получил название конского хвоста, caucla equina. Внутреннее строение. На поперечном разрезе спинного мозга серое вещество, которое окружает центральный канал, canalis centralis, представлено парными передними, cornu anterius, задними, cornu posterius, и боковыми рогами (С8—L2), cornu lateralis (см. рис. 2.6), которые в действительности представляют собой непрерывные столбы (columnae griseae), идущие вдоль спинного мозга. Рога серого вещества обеих половин спинного мозга соединяются с помощью передней серой спайки, conunissura grisea anterior, и задней серой спайки, commissura griseaposterior. Спереди от commissura grisea anterior располагается передняя белая спайка, commissura alba anterior. В сером веществе спинного мозга тела нервных клеток, сходных по строению и функции, объединяются, образуя ядра. В переднем роге находится группа так называемых двигательных ядер, которые состоят преимущественно из мотонейронов. Различают крупные альфа-мотонейроны (диаметр тела 35-70 мкм) и рассеянные среди них более мелкие гамма-мотонейроны (диаметр гела 15—35 мкм). Аксоны мотонейронов покидают спинной мозг в составе передних корешков, затем в составе смешанных спинномозговых нервов направляются к соматическим мышцам. Описано 5 ядер переднего рога, которые соответственно положению получили названия: переднемедиальное, заднемедиальное, перед нелатеральное, заднелатеральное и центральное. Ядра заднего рога образованы главным образом ассоциативными нейронами (интернейронами) и включают: • студенистое вещество, substantia gelatinosa, — образовано телами ассоциативных нейронов переднего спинноталамического пути; • собственные ядра, nuclei proprii, — образованы телами ассоциативных нейронов латерального спинноталамического пути; • грудное ядро, nucleus thoracicus (столб Кларка— Штиллинга, columna Stilling—Clarke) (С8—L2), — образовано телами ассоциативных нейронов заднего спинно-мозжечкового пути. В промежуточной зоне серого вещества находятся: • латеральное промежуточное ядро, nucleus intermediolateralis (C8-L2), — центр симпатической нервной системы; • медиальное промежуточное ядро, nucleus intermediomedia- lis, — образовано телами ассоциативных нейронов переднего спинно-мозжечкового пути. Белое вещество спинного мозга окружает серое вещество и разделяется на симметричные задние, боковые и передние канатики: funiculus anterior, funiculus lateralis и funiculus posterior (см. рис. 2.6). Оно образовано миелиновыми нервными волокнами, которые группируются в проводящие пути (рис.2.8). Различают
Рис. 2.8. Топография проводящих путей спинного мозга: 1— fasciculus gracilis; 2 — fasciculus cuneatus; 3 — fasciculus dorsolateralis; 4 — tracfus spinocerebellaris posterior; 5 — tractus spinothalamicus lateralis; 6 — tractus spinocerebel- laris anterior; 7 — tractus spinoolivaris; 8 — spinotectalis; 9 — tractus spinothalamicus anterior; 10— fasciculus longitudinalis medialis; 11 — tractus corticospinalis anterior; 12 — tractus tectospinalis; 13 — tractus reticulospinalis; 14 — tractus vestibulospinalis; 15 — tractus reticulospinalis; 16 — tractus rubrospinalis; 17 — tractus corticospinalis lateralis; 18 — fasciculus interfascicularis; 19 — fasciculus septomarginalis
нисходящие (двигательные, эфферентные) и восходящие (чувствительные, афферентные) пути. Двигательные пути, которые начинаются из коры больших полушарий, являются проводниками сознательных двигательных импульсов на скелетную мускулатуру. Они входят в состав так называемой пирамидной системы, которая обеспечивает всю совокупность произвольных движений организма. Двигательные пути, которые начинаются из подкорковых ядер головного мозга, являются проводниками бессознательных двигательных импульсов на скелетную мускулатуру. Они входят в состав экстрапирамидной системы. Восходящие пути в зависимости от вида чувствительности, проводниками которой они являются, подразделяются на пути экстероцептив- ной (тактильной, болевой, температурной) и проприоцептивной (мышечносуставное чувство) чувствительности. При этом аналогично проводникам двигательных импульсов они подразделяются на пути сознательных и бессознательных ощущений. Нисходящие пути: Проводящие пути пирамидной системы: • латеральный корково-спинномозговой (пирамидный) путь, tr. corticospinalis (pyramidalis) lateralis (17); • передний корково-спинномозговой (пирамидный) путь, tr. corticospinalis (pyramidalis) anterior (11). Проводящие пути экстрапирамидной системы: • красноядерно-спинномозговой путь (Монаков), tr. rubrospi- nalis (16); • покрышечно-спинномозговой путь, tr. tectospinalis (12); • преддверно-спинномозговой путь, tr. vestibulospinalis (14); • оливоспинномозговой путь, tr. olivospinalis; • ретикулярно-спинномозговой путь, tr. reticulospinalis (13, 15); • медиальный продольный пучок, fasciculus longitudinalis те- dialis (10). Нисходящие пути: • боковой спинно-таламический пучь, tr. spinothalamicus lateralis, — проводящий путь болевой и температурной чувствительности коркового направления (5); • передний спинно-таламический путь, tr. spinothalamicus anterior, — проводящий путь тактильной чувствительности коркового направления (9); • тонкий пучок (Голля), fasciculus gracilis, — проводящий путь проприоцептивной чувствительности коркового направления (от нижних конечностей и нижней половины туловища) (1); • клиновидный пучок (Бурдаха), fasciculus cuneatus, — Проводящий путь проприоцептивной чувствительности коркового направления (от верхних конечностей и верхней половины туловища) (2); • передний спинно-мозжечковый путь, rr. spinocerebellaris ventralis (anterior), — проводящий путь проприоцептивной чувствительности к мозжечку (6); • задний спинно-мозжечковый путь, tr. spinocerebellaris dorsalis (posterior), — проводящий путь проприоцептивной чувствительности к мозжечку (4). Оболочки спинного мозга. Спинной мозг окружен твердой, dura mater spinalis, паутинной, arachnoidea spinalis, и мягкой, pia mater spinalis, оболочками. Твердая мозговая оболочка образована плотной волокнистой соединительной тканью и образует просторный футляр, который простирается от уровня большого затылочного отверстия до II крестцового позвонка. Между твердой оболочкой спинного мозга и позвонками находится эпидуральное пространство, cavitas epiduralis, заполненное рыхлой волокнистой соединительной тканью и жировой клетчаткой, содержащей большое число лимфатических сосудов и густое венозное сплетение. Внутренняя поверхность твердой мозговой оболочки отделяется от паутинной оболочки капиллярным субдуральным пространством, spatium subdurale. Тонкая бессосуди- стая паутинная оболочка лежит между твердой и мягкой оболочками и отделена от последней субарахноидальным пространством, cavitas subarachnoidalis, в котором циркулирует спинномозговая жидкость. Эта полость с помощью зубчатой связки, liga- mentum denticulatum, делится на переднюю и заднюю части. Мягкая мозговая оболочка плотно прилегает к спинному мозгу и сращена с ним. Она состоит из двух листков, между которыми находится так называемое интерпиальное пространство, где располагается сеть кровеносных сосудов. ОБЩИЙ ОБЗОР ГОЛОВНОГО МОЗГА
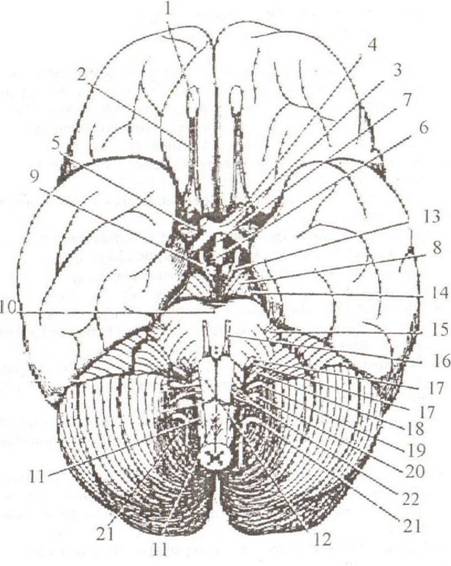
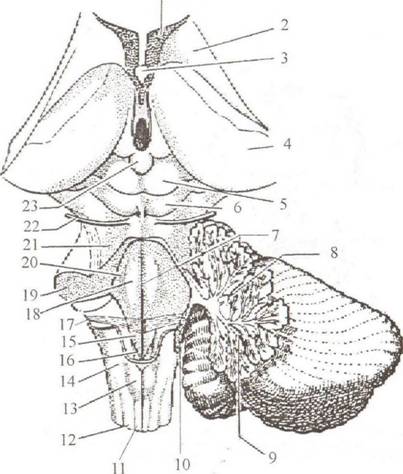
Основные части головного мозга: большой мозг, ствол и малый мозг (мозжечок). Полушария большого мозга разделены глубокой вертикальной щепъю, Jissura longitudinalis cerebri. Поперечная щель, fissura transversa cerebri, отделяет большой мозг от пижерасположенного мозжечка, cerebellum. На поверхности полушарий определяются борозды, sulci cerebri, между которыми расположены извилины, gyri cerebri. Вентральная (нижняя) поверхность называется основанием мозга, basis cerebri. По направлению от лобного полюса к затылочному здесь можно увидеть следующие образования (рис. 2.9): • Обонятельная луковица, bulbus olfactorius (I). • Обонятельный тракт, traclus olfactorius (2). • Обонятельный треугольник, trigonum olfactorium, который ограничен наружной и внутренней обонятельными полосками, striae olfactoriae medial is et lateralis. • Перекрест зрительных нервов, chiasma opticum (3). Спереди от перекреста находятся зрительные нервы, opticus, сзади — зрительные тракты, tractus opticus. • Переднее продырявленное вещество, substantia perforata anterior (5), расположенное по бокам от перекреста и зрительных трактов. • Серый бугор, tuber cinereum (6), который вытягивается в отросток, напоминающий воронку, infundibulum', с его помощью основание мозга соединяется с гипофизом, hypophysis. • Ножки мозга, crus cerebri (8), находятся справа и слева от серого бугра. • Сосцевидные тела, corpora mamillaria (9), лежат позади серого бугра. • Межножковая ямка, fossa interpeduncularis, занимает пространство между ножками мозга и кзади от сосцевидных тел. • Заднее продырявленное вещество, substantia perforata posterior, образует дно межножковой ямки.
Рис. 2.9. Основание головного мозга: 1— bulbils olfactorius; 2 — iractus olfactorius;3 — chiasm a optician; 4 — a. opticus; 5 — substantia perforata anterior; 6 — tuber cinereum; 7 — infundibulum, S — crus cerebri; 9 — corpora mamillaria; 10 — pons Varoli; II — medulla oblongata; 12 — pyramis; 13 — n. oculomotorius (iii); 14 — n. trochlearis (IV); 15 — n. trigeminus (V); 16 — n. abducens (vi); 17— n. facialis (vii); 18 — n. vestibulocochlearis ( viii); 19 — n. glossopharyngeus (ix); 20 — n. vagus (X); 21 — n. accessorius (xi); 22 — n. hypoglossus (xii) • Мост, pons Varoli (10), с расположенной посередине срединной бороздой, sulcus basilaris; кнаружи мост суживается и затем, направляясь в стороны и назад, погружается в мозжечок, формируя его средние ножки, peduneulus cerebellaris medius. • Продолговатый мозг, medulla oblongata (II), лежит позади моста. По средней линии продолговатого мозга проходит передняя срединная щель, fissura mediana anterior, ограниченная с каждой стороны белым валиком, пирамидой, ру- ratnis. Кнаружи от пирамиды лежит продолговатое возвышение, олива, oliva. На основании мозга можно увидеть корешки 12 пар черепных нервов: • Обонятельный нерв, п. olfactorius (I). Первую пару черепных нервов составляют центральные отростки обонятельных клеток, расположенных в слизистой оболочке носовой полости, — это так называемые обонятельные нити,,№ ol- factoria. Они проникают в полость черепа через продырявленную пластинку решетчатой кости и подходят к обонятельной луковице. • Зрительный нерв, //. opticus (II), образует с одноименным нервом противоположной стороны зрительный перекрест, chiasma opticum, и далее продолжается под названием tractus opticus. • Глазодвигательный нерв, п. oculomotorius (III), выходит на внутренней поверхности ножек мозга из одноименной борозды. • Блоковый нерв, п. trochleans (IV), выходит из верхнего мозгового паруса и появляется на основании снаружи от ножки мозга. • Тройничный нерв, п. trigeminus (V), лежит у переднего края варолиева моста, на границе со средними ножками мозжечка. Появляется двумя корешками: тонким — двигательным и толстым — чувствительным. • Отводящий нерв, п. abducens (VI), выходит на границе заднего края моста и пирамиды продолговатою мозга. • Лицевой нерв, п. facialis (VII), находится между задним краем моста и оливой, в так называемом мосто-мозжечковом углу. • Преддверно-улитковый нерв, п. veslibulocochlearis (VIII), лежит в мосто-мозжечковом углу снаружи от лицевого нерва. • Языкоглоточный нерв, п. glossopharyngeus (IX), 5-6 корешков этого нерва находятся позади олив. • Блуждающий нерв, п. vagus (X), 10—12 корешков этого нерва лежат позади олив ниже предыдущей пары. • Добавочный нерв, accessorius (XI), выхолит многими корешками на боковой поверхности продолговатого мозга и шейной части спинного мозга. • Подъязычный нерв, n. hypoglossus (XII), появляется на основании мозга между пирамидой и оливой. На сагиттальном разреземозга определяются (рис. 2.10):
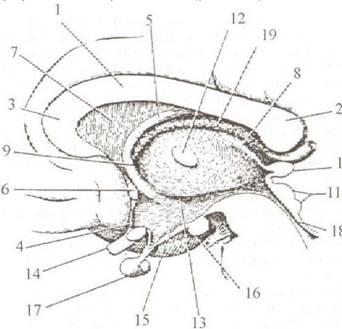
Рис. 2.10. Фрагмент сагиттального разреза головного мозга: 1 — corpus callosum; 2 — splenium; 3 — genu; 4 — lamina ter fornix; 6 — conunissura cerebri anterior; 7 — septum pellu thalamus; 9 — foramen interven- triculare; 10 — epiphysis; 11— (lamina quadrigemina); 12 — adhesio intert- halamica; 13 hypothalamicus; 14 — cliiasma opticum; 15 — tuber cinereum. mamillare; 17 — hypophysis; 18 — aquaeductus cerebri plexux chorioideus • Мозолистое тело, corpus callosum (1), мал которым располагается медиальная поверхность полушария большого мозга и мозжечка. Среднюю часть мозолистого тела называют стволом, truncus corporis callosi, кзади мозолистое тело утолщается, образуя валик, splenium, а спереди изгибается дугою вниз и формирует колено, genu, которое, суживаясь, переходит в клюв, rostrum. Клюв продолжается в короткую и тонкую мозговую пластинку, lamina rostralis, к которой затем примыкает конечная пластинка, lamina terminalis, направляющаяся к передней поверхности зрительного перекреста. • Свод, fornix (5), находится под мозолистым телом, от которого отделяется на границе валика и ствола и в виде выпуклой вперед дуги проникает в глубину мозгового вещества позади поперечно перерезанного белого пучка волокон, передней мозговой спайки, commissura cerebri anterior. • Прозрачная перегородка, septumpellucidum (7), лежит между сводом и мозолистым телом. • Зрительный бугор, thalamus (8), находится под сводом. • Межжелудочковое отверстие, foramen interventriculare (9), располагается между передним концом зрительного бугра и сводом. • Эпифиз, epiphysis (corpus pineale) (10), находится у заднего конца зрительного бугра, под валиком мозолистого тела. • Шишковидное углубление, recessuspinealis, — щель, проникающая спереди в corpus pineale. • Задняя мозговая спайка, commissura cerebri posterior, расположена ниже шишковидного углубления. • Пластинка четверохолмия, lamina tecti (lamina quadrigemina) (II) , — дорсальная часть среднего мозга. • Верхипп мозговой парус, velum medullare superius, располагается сразу под четверохолмием. • Мозжечок, cerebellum. • Межбугорное сращение (поперечный разрез), adhesio intert- halamica (12), лежит на медиальной поверхности зрительного бугра, позади межжелудочкового отверстия. • Подбугорная борозда, sulcus hypothalamicus (13), расположена под межбугорным сращением и ограничивает нижележащую гипоталамическую область. Здесь на сагиттальном разрезе видны уже упомянутые при описании основания мозга образования: • Зрительный перекрест, chiasma opticum (14). • Воронка, infundibulum. • Серый бугор, tuber cinereum (15). • Сосцевидное тело, corpus mamillare (16). • Гипофиз, hypophysis (17). • Заднее продырявленное вещество, substantia perforata posterior. • Межножковая ямка, fossa interpeduncularis. • Ножка мозга, crus cerebri. • Мост, pons. • Продолговатый мозг, medulla oblongata. • Водопровод мозга, aquaeductus cerebri (Sylvii) (18), к которому дальше примыкает IV желудочек, лежащий под мозжечком.
РОМБОВИДНЫЙ МОЗГ
В состав ромбовидного мозга входят собственно задний мозг, metencephalon, и продолговатый мозг, myelencephalon, а его полостью является четвертый желудочек, ventriculus quartus. Metencephalon включает мост, pons, и мозжечок, cerebellum.
ПРОДОЛГОВАТЫЙ МОЗГ
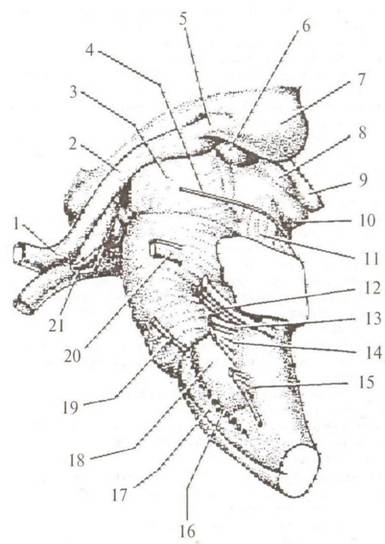
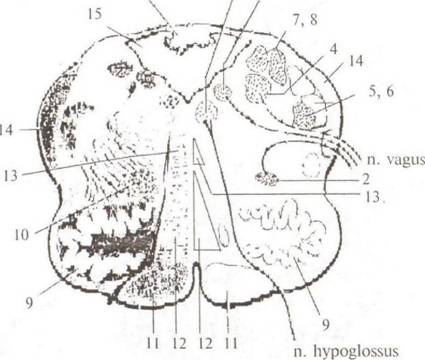
Внешнее строение. В продолговатом мозге выделяют вентральную, дорсальную и боковые поверхности. На вентральной поверхности верхней границей продолговатого мозга является задний край варолиева моста. За нижнюю границу принимают место выхода корешков первой пары шейных спинномозговых нервов. Рельеф вентральной поверхности определяют следующие образования (рис. 2.11): • Передняя срединная шель, fissura mediana anterior. • Пирамида, pyramis (17), которая определяется с каждой стороны от передней срединной щели и образована волокнами корково-спинномозгового пути. Большая часть
Рис. 2.11. Стволовая часть мозга (вентролатеральная поверхность): 1 — chiasma opticum; 2 — tractus opticas; 3 — eras cerebi; 4 — n. trochlearis; 5 — corpus geniculatum laterale; 6 — corpus geniculatum mediate; 7 — pulvinar; 8 — collacalas superior; 9 — corpus pineate; 10 — colliculus inferior; 11 — pedunculus cerebellaris inferior; 12 — n. faciacis et n. vestibulocochlearis; 13 — n. gtossofaryngeus; 14 — n. vagus; 15 — n. accessorius; 16 — n. hypoglossus; 17 — pyramis; 18 — oliva; 19 — n. abducens; 20 — n. trigeminus; 21 — infundibulum волокон этого пути переходит среднюю линию (decussatio pyramidum) и далее проходит в боковом канатике противоположной стороны спинного мозга. • Олива, oliva (18), находится снаружи от пирамиды. • Корешки подъязычного нерва (16), расположенные между пирамидой и оливой. • Передняя боковая борозда, sulcus lateralis anterior, где локализуются корешки IX, X и XI пар черепных нервов. Верхней границей продолговатого мозга на дорсальной поверхности являются медуллярные полоски ромбовидной ямки, striae medullares fossae rhomboideae. Рельеф дорсальной поверхности определяют следующие образования (рис. 2.12): • Задняя срединная борозда, sulcus medianus posterior (7), которая сверху замыкается тонким мозговым листком — задвижкой, obex. Под задвижкой идущий в глубине центральный канал переходит в четвертый желудочек. • Задняя промежуточная борозда, sulcus intermedius posterior (11), которая разделяет восходящие из спинного мозга нежный и клиновидный пучки, fasciculi gracilis (11) и сипеа- tus (12). • Бугорки тонкого и клиновидного ядер, tuberculi gracile (13) и cuneatum (14), которыми заканчиваются одноименные пучки. • Задняя боковая борозда, sulcus lateralis posterior, ограничивает снаружи клиновидный пучок. Между передней и задней боковыми бороздами располагается боковая часть продолговатого мозга, которая продолжается в нижние ножки мозжечка. Внутреннее строение (рис. 2.13). Серое вещество продолговатого мозга представлено следующими образованиями: • Ядра черепных нервов: ■ Двигательные ядра подъязычного (XII) и добавочного (XI) нервов. ■ Двигательные и чувствительные соматические, а также парасимпатические ядра блуждающего (X) и языкоглоточного (IX) нервов. ■ Чувствительное ядро тройничного нерва, которое распространяется на всем протяжении продолговатого мозга, опускаясь в верхние шейные сегменты спинного мозга.
Рис. 2.12. Стволовая часть мозга (дорсальная поверхность): 1 —cornu frontale; 2 —nucleus caudatus; 3 —columna fornicis; 4 — thalamus: 5 — colliculus superior; 6 — colliculus inferior; 7—sulcus medianus posterior; 8, 9 — cerebellum; 10 — pedunculus cerebellaris inferior; II — fasciculus gracilis; 12 — fasciculus cuneatus; 13 — tuberculum gracile; 14 — tuherculum cuneatum; 15 — trigonum nervi hypoglossi; 16 — trigonum nervi vagi; 17 — striae medullares; 18 — eminentia medialis; 19 — pedunculus cerebellaris medius; 20 — fovea superior; 21 — pedunculus cerebellaris superior; 22 — n. trochlean's; 23 — corpus pineale
Рис. 2.13. Поперечный разрез продолговатого мозга: 1 — nucleus motorius ncrx’i hypoglossi; 2 — nucleus ambiguus; 3 — nucleus dorsalis nervi vagi; 4 — nucleus solitarius; 5,6 — nucleus el Iraclus spinalis nervi trigemini; 7, 8 — nuclei vestibulares; 9 — nucleus olivaris inferior; 10 — formatio reticularis; 11— pyramis; 12 — lemniscus medialis; 13 — fasciculus longitudinalis medialis; 14 — pedunculus ccrebcllaris inferior; 15 — ventriculus quartus 16 — plexus chorioideus
• Ядерный комплекс оливы, который включает основное нижнее оливное ядро, nucleus olivaris inferior, и добавочные (медиальное и заднее) оливные ядра, nucleus olivaris accessorius medialis и posterior. Эти ядра относятся к экстрапирамидной системе и имеют связи с мозжечком и спинным мозгом: ir. olivospinalis, tr. spinoolivaris, tr. olivocerebellaris. • Ядра тонкого (Голля) и клиновидного (Бурдаха) пучков, nucleus gracilis и nucleus cuneatus, расположенные в одноименных бугорках дорсальной поверхности продолговатого мозга, образованы телами ассоциативных нейронов восходящих проводящих путей проприоцептивной чувствительности коркового направления (пути Голля и Бурдаха). Аксоны вторых нейронов, выходящие из указанных ядер, переходят на противоположную сторону продолговатого мозга, образуя так называемую медиальную петлю мозгового ствола. Перекрест нервных волокон в дорсальной части продолговатого мозга получает название перекреста медиальных петель, или чувствительного перекреста, decussatio lemniscorum media Hum (decussatio sensoria). • Ретикулярная формация, formatio reticularis. Состоит из отдельных групп клеток и многочисленных нервных волокон, которые проходят в различных направлениях, устанавливая синаптические связи со всеми отделами ЦНС. Осуществляет регуляцию и интеграцию разнообразных вегетативно-висцеральных и сенсомо- торных реакций. Скопления клеток в ретикулярной формации продолговатого мозга образуют сосуд од вигательный (вазомоторный) и дыхательный центры. Вазомоторный центр является сложным образованием, включает в себя депрессорный и прессорный центры, регулирующие артериальное давление; акселера- торный и ингибиторный центры, регулирующие частоту сердечных сокращений. Дыхательный центр расположен в боковых отделах ретикулярной формации и включает центр вдоха и выдоха. Белое вещество продолговатого мозга включает проводящие пути восходящего и нисходящего направлений: Нисходящие проводящие пути: • Tr. corticospinalis (pyramidalis) занимает вентральное положение, с каждой стороны от передней срединной щели, на границе со спинным мозгом подвергается неполному перекресту, который получает название перекреста пирамид или моторного перекреста, decussatio pyramidum (decussatio motoria). Большая часть волокон переходит на противоположную сторону и проходит в боковом канатике спинного мозга (tr. corticospinalis lateralis). Меньшая часть волокон продолжается в переднем канатике своей стороны (tr. Corticospinalis anterior). • Проводящие пути экстрапирамидной системы: tr. rubrospi- nalis, tr. tectospinalis, tr. vestibulospinalis, tr. reticulospinalis, tr. olivospinalis, fasciculus longitudinalis nicdialis. Восходящие проводящие пути: • Lemniscus medialis, медиальная петля, образована аксонами вторых нейронов проводящих путей проприоцептивной чувствительности (Голля и Бурдаха) противоположной стороны. • Tr. spinothalamicus, спинно-таламический путь — проводящий путь экстероцептивпой чувствительности коркового направления. В области мозгового ствола получает название спинномозговой петли, lemniscus spinalis. На всем протяжении до зрительного бугра она сопровождает волокна медиальной петли, располагаясь снаружи от проводников п роп р и оце пти вн о й чу вств ите л ьн ости. • 7г. spinocerebellaris ventralis (Gowers), передний спинномозжечковый путь — проводящий путь проприоцептивной чувствительности. Располагается в латеральной части продолговатого мозга между оливой и нижними ножками мозжечка. Волокна заднего спинно-мозжечкового пути (Flech- sig) покидают продолговатый мозг и в составе нижних ножек мозжечка направляются в кору червя.
МОСТ
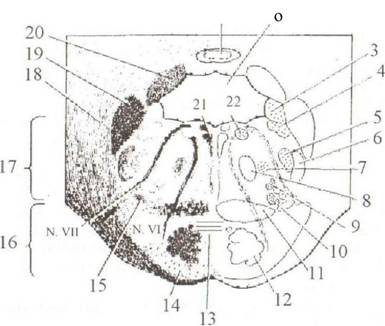
Внешнее строение. Мост, pons, расположен между ножками мозга сверху и спереди и продолговатым мозгом снизу и сзади (см. рис. 2.9). На вентральной поверхности моста находятся следующие образования: • Бульбарно-мостовая борозда, sulcus bulbopontinus, расположенная между продолговатым мозгом и нижним краем моста, где находятся корешки отводящего (VI) нерва. • Мостомозжечковый угол (треугольник), trigonum pontocere- bellare, который лежит между мостом, продолговатым мозгом и мозжечком. Здесь локализуются волокна лицевого (VII) и преддверно улиткового (VIII) нервов. • Тройнично-лицевая линия, tinea trigeminofacialis, соединяет места выхода корешков тройничного (V) и лицевого (VII) нервов и является боковой границей моста. • Средние ножки мозжечка, pedunculi cerebellaris medii, лежат снаружи от тройнично-лицевой линии. • Базилярная борозда, sulcus basilaris, в которой лежит одноименная артерия {a. basilaris). Дорсальная часть моста принимает участие в образовании ромбовидной ямки, которая является дном IV желудочка (см. рис. 2.12). Здесь на границе с продолговатым мозгом определяются лицевой бугорок, colliculus facialis, и мозговые полоски, striae medul/ares, которые являются частью слухового пути. Внутреннее строение (рис. 2.14). Волокна трапециевидного тела, corpus trapezoideum, разделяют мост на вентральную, pars basilaris, и дорсальную, pars dorsalis (tegmentum), части. В pars dorsalis ponds серое вещество представлено: Ядрами черепных нервов: • тройничного (V) — двигательное и чувствительное соматические ядра; • отводящего (VI) — двигательное соматическое ядро; • лицевого (VII) — двигательное и чувствительное соматические и вегетативное парасимпатическое ядра; • преддверно-улиткового (VIII) — чувствительные ядра. При этом наиболее дорсальное положение занимает двигательное ядро отводящего нерва. Ядро лицевого нерва лежит значительно более вентрально. Волокна, выходящие из ядра лицевого нерва, направляются кверху, огибают ядро отводящего, образуя колено лицевого нерва, genu п. facialis, и снова поворачивают в вентральном направлении, выходя в мосто-мозжечковом углу. Ядра тройничного нерва располагаются снаружи и выше ядра VII пары. Чувствительные ядра преддверно-улиткового нерва находятся на границе с продолговатым мозгом снаружи от corpus trapezoideum. Серое вещество покрышки моста также образовано многочисленными ядрами ретикулярной формации ствола. Белое вещество — система восходящих и нисходящих проводящих путей. Восходящие пути составляют: • медиальная петля, lemniscus medialis\ • спинномозговая петля, lemniscus spinalis;
Рис. 2.14. Поперечный разрез мости: 1—cerebellum; 2 — ventriculus quartus; 3, 4 — nuclei vestibulares; 5, 6 — nucleus el trac- tus spinalis n. trigemini; 7—nucleus reticularis caudalis pontis; 8 — tractus tegmentalis centralis; 9 — nucleus nervifacialis; 10— nucleus otivaris superior; 11 — lemniscus medialis; 12 — tractus corticospinalis; 13 — corpus trapezoideum; 14 —nuclei proprii pontis; 15 —lemniscus lateralis; 16 — pars ventralis pontis; 17 — pars dorsalis pontis; 18, 19, 20 — pedunculli ccrehellares medius, inferior et superior; 21 — fasciculus longituclinalis medialis; 22 — nucleus nen' abducentis
• латеральная петля, lemniscus lateralis (является продолжением волокон трапециевидного тела и составляет часть слухового пути). Нисходящие волокна образованы проводящими путями эк- страпирамидной системы (fasciculus longituclinalis medialis, tr. tec- tospinalis, tr. reticulospinalis и др.). В pars ventralis pontis серое вещество представлено собственными ядрами моста, nuclei proprii pontis, которые образованы телами ассоциативных нейронов проводящего пути, соединяющего кору больших полушарий с корой мозжечка (tr. cortico-pontocerebellaris). Белое вещество вентральной части моста представлено продольно и поперечно идущими волокнами, fibrae longitudinciles и fibrae transversae. Продольные волокна включают две составные части пирамидного пути (/г. corticospinalis и tr. corticobulbciris) и корково-мостовые волокна, которые соединяют кору лобной, затылочной и височной долей с собственными ядрами моста {fibrae corticopontinae). Поперечно идущие пучки волокон начинаются от собственных ядер моста и направляются в среднюю мозжечковую ножку противоположной стороны (Jibrae ponto-ce- rebellares). Поперечные волокна, которые локализуются вентрально от пирамидных путей, называются поверхностными, fibrae transversae supeificiales, а лежащие дорсально — глубокими, fibrae transversae profundae.
МОЗЖЕЧОК
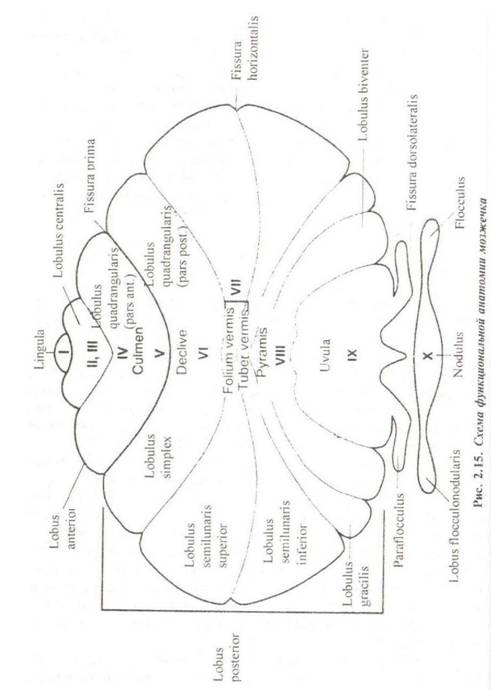
Внешнее строение. Мозжечок, cerebellum, состоит из непарной срединной части, которая называется червь, vermis, и боковых частей — полушарий мозжечка, hemispheria cerebelli. В мозжечке различают верхнюю и нижнюю поверхности, facies superior и inferior, передний и задний края, margo anterior и posterior, на которых находятся одноименные вырезки, ineisura cerebelli anterior и posterior, передние, задние и боковые углы, anguli anteriores, posteriores и latera/es. Посередине нижней поверхности находится широкая борозда — долинка мозжечка, valecula cerebelli, в которой помещается продолговатый мозг. Рельеф мозжечка представлен многочисленными бороздками, sulci cerebelli, которые отделяют друг от друга узкие извилины, gyri cerebelli. На границе верхней и нижней поверхностей мозжечка проходит глубокая поперечная борозда, sulcus horizonta/is. Полушария разделяются постоянными глубокими бороздами на три доли: lobus anterior, lobus posterior и lobus flocculonodularis, которым соответствуют определенные участки червя. Каждая доля разделена на дольки, которые обозначаются как классическими названиями, так и широко распространенной нумерацией по Ларселу (римские цифры) (рис. 2.15). В ходе филогенеза происходит постепенное усложнение строения мозжечка параллельно развитию полушарий большого мозга.
В соответствии с филогенетическим возрастом различают три части мозжечка: • старая часть, archaeocerebellum, к которой относятся клочок, узелок и язычок червя; • древняя часть, paleocerebellum, которая состоит из язычка, центральной дольки, вершины и пирамиды червя, а также крыла центральной дольки и четырехугольной дольки (передняя часть) полушарий; в филогенезе появляется после старой части; • новая часть, neocerebellum, представленная скатом, листком и бугром червя, четырехугольной (задняя часть), верхней и нижней полулунными дольками и миндалиной полушарий мозжечка; в филогенезе появляется позднее остальных частей. Эта классификация соответствует подразделению органа на отделы в соответствии с проекцией афферентных волокон мозжечкового направления. Так, старая часть называется vestibuloce- rebellum, так как здесь заканчиваются пути из вестибулярных ядер. Древняя часть получает название spinocerebellum, так как в нее приходят пути из спинного мозга. Наконец, новая часть, pontocerebellum, получает афферентные волокна из коры больших полушарий. Внутреннее строение. Вся наружная поверхность органа покрыта серым веществом, cortex cerebelli. Кора мозжечка имеет трехслойное строение (рис. 2.16): • Молекулярный слой, stratum moleculare, — наружный слой коры. Состоит преимущественно из ветвящихся дендритов и аксонов, а также небольшого количества тел звездчатых и корзинчатых нейронов. • Слой грушевидных нейронов или слой клеток Пуркинье. stratum neurium piriformium, — средний слой коры. У человека в нем насчитывается до 15 млн клеток. Дендриты грушевидных нейронов ветвятся в молекулярном слое, большая часть аксонов направляется к ядрам мозжечка, а меньшее их число — к вестибулярному ядру Дейтерса. • Зернистый слой, stratum granu/osum, — внутренний слой коры. Он состоит из многочисленных мелких клеток- зерен. Их аксоны направляются в молекулярный слой, где Т-образно делятся, образуя ветви, направленные вдоль
Рис. 2.16. Строение коры мозжечка: 1 — молекулярный слой; 2 — слой клеток Пуркинье; 3 — зернистый слой поверхности коры мозжечка (параллельные волокна). В зернистом слое также находятся более крупные клетки Гольджи, дендриты которых распространяются в молекулярном слое, а аксоны подходят к клеткам- зернам. В кору мозжечка входят два типа волокон: • Лазящие (лиановидные) волокна оплетают дендриты клеток Пуркинье. Эти волокна берут начало в ядрах нижней оливы. • Мшистые волокна отдают множество коллатералей и заканчиваются на клетках-зернах. Такого типа волокна характерны для всех остальных афферентных проекций мозжечка. В толше мозжечка серое вещество представлено ядрами: • зубчатое ядро, nucleus dentatus, имеет вид изогнутой пластинки с медиально-расположенными воротами; • пробковидное ядро, nucleus emboliformis, расположено спереди от ворот зубчатого ядра; • шаровидное ядро, nucleus globosus. находится с медиальной стороны от зубчатого ядра; • ядро шатра, nucleusfastigii, — самое внутреннее из ядер мозжечка. Лиановидные волокна образуют многочисленные возбуждающие синапсы на дендритах клеток Пуркинье. Мшистые волокна возбуждают клетки-зерна, которые через параллельные волокна оказывают возбуждающее действие на все остальные нейроны. Действие последних, однако, всегда является тормозным: клетки Гольджи тормозят клетки-зерна, а разряды клеток Пуркинье приводят к торможению нейронов ядер мозжечка. Таким образом, за исключением клеток-зерен, все нейроны, тела которых расположены в коре мозжечка, выполняют тормозные функции. Ни в одном другом отделе центральной нервной системы нет такого преобладания торможения над возбуждением. Нейроны Пуркинье обладают некоторой активностью покоя, обусловливающей тоническое торможение ядер мозжечка. При увеличении активности клеток Пуркинье торможение ядер мозжечка усиливается, торможение же нейронов Пуркинье сопровождается растормаживал нем этих ядер. Кора червя и полушарий мозжечка характеризуется сомато- топической организацией: в передних отделах полушарий представлены верхние конечности, а в задних отделах — нижние; в передних отделах коры червя — голова и шея, а в задних отделах — туловище. Проксимальные отделы конечностей проецируются медиальнее, дистальные — латеральнее; полушария ответственны за координацию движений конечностей, червь — туловища (рис. 2.17).
Рис. 2.17. Соматотопическая проекция в коре червя и полушариях мозжечка Белое вещество мозжечка получило название мозгового тела, corpus mcdullare, от которого отходят сначала более крупные мозговые пластинки белог о вещества, laminae medullares, которые, в свою очередь, отдают вторичные мозговые листки. Последние дают начало еще более мелким листочкам, которые покрыты серым веществом и образуют извилины мозжечка, gyri cerebelli. Картина расположения белого вещества, видимая на сагиттальном разрезе, вследствие внешнего сходства с ветвлением дерева называется arbor medullaris. Белое вещество полушарий соединяется с соседними частями мозга посредством ножек мозжечка. Средние мозжечковые ножки, peduncu/i cerebellares medii. направляются в варолиев мост. Они содержат волокна мостомозжечкового пути, tr. ponto- cerebellaris. Верхние мозжечковые ножки, pedunculi cerebellares su- periores, связывают мозжечок со средним мозгом. Между ними находится верхний мозговой парус. В верхних ножках проходят волокна переднего спинномозжечкового пути, tr. spinocerebella- ris ventralis, которые совершают перекрест в области верхнего мозгового паруса. В верхних ножках также локализуются крас- ноядерно- зубчатые волокна, fibrae dentatae rubrales, которые соединяют зубчатое ядро мозжечка с красным ядром покрышки ножек мозга. Нижние мозжечковые ножки, pedunculi cerebellares in- feriores, идут к продолговатому мозгу. Здесь проходят задний спинно-мозжечковый путь, tr. spinocerebellaris dorsalis, оливомозжечковый путь, tr. olivocerebellaris, задние наружные дугообразные волокна, fibrae arcuatae externae posteriores, которые начинаются от добавочного клиновидного ядра и формируют tr. сипео- cerebellaris — проводник проприоцептивных импульсов от верхних конечностей к мозжечку. Этот путь замещает задний спинно-мозжечковый путь в сегментах спинного мозга, расположенных выше С8. |
||
|
|
Последнее изменение этой страницы: 2018-05-10; просмотров: 562. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |