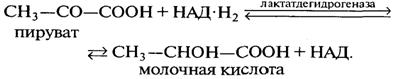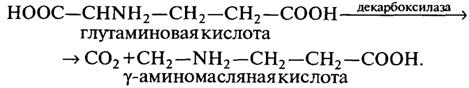|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
МЕТАБОЛИЗМ И БИОХИМИЧЕСКИЕ СВОЙСТВА МОЛОЧНОКИСЛЫХ БАКТЕРИЙМолочнокислые бактерии являются постоянными спутниками муки, дрожжей, молочных продуктов и др. Они оказывают влияние на вкус, аромат и усвояемость хлеба. Содержание молочнокислых бактерий в муке зависит от ее сорта. По данным В. М. Богдановой, содержание микроорганизмов в муке (% к об-семененности зерна): высшего сорта — 9—14; первого сорта — 11—32; второго сорта — 25—56; отрубях — до 50. Среди общего числа микроорганизмов определенная доля принадлежит и молочнокислым бактериям. Свойство молочнокислых бактерий образовывать молочную кислоту объединяет их в отдельную группу микроорганизмов, которая по практической значимости занимает одно из первых мест. Эти бактерии участвуют в многочисленных биохимических процессах. Они представляют собой одноклеточные микробы размером от 2 до 10 мкм, шарообразной и палочковидной форм, неподвижные, неспособные к образованию спор, грамположи-тельны и не чувствительны к кислороду. Рациональный температурный интервал для роста бактериальных клеток 45—47 °С; для кислотообразования 52—54 °С. Бактериальные клетки делятся через каждые 60 мин, а дрожжевые — каждые 1,5—2 ч. По отношению к температуре бактерии делят на 3 группы: психрофилы — температурный оптимум 10 °С, минимальная температура около 0 и максимальная — 30 °С; мезофилы — соответственно 30; 0 и 42 °С; термофилы — наиболее благоприятная температура для развития 50—70 °С; максимальная для жизнедеятельности — 75 °С. К молочнокислым бактериям относятся представители родов Leuconostoc, Streptococcus, Lactobacillus. Бактерии рода Leuconostoc. К этому роду относятся гетероферментативные молочнокислые бактерии, продуцирующие наряду с молочной кислотой большое количество летучих соединений (уксусная кислота, этанол, СО2 и др.); для ак-  тивного роста нуждаются в витаминах. Клетки неподвижны, имеют вид спаренных удлиненных кокков диаметром 0,9—1,2мкм. Бактерии рода Leuconostoc на твердых средах с сахарозой образуют колонии в виде капель слизи, без сахарозы — колонии правильной круглой формы, пастообразной консистенции, с гладкой блестящей поверхностью грязно-белого цвета. Бактерии рода Streptococcus продуцируют только молочную кислоту. Бактерии рода Lactobacillus. Представителями рода Lactobacillus являются Lactobacillus plantarum, Lactobacillus fermenti, Lactobacillus brevis, Lactobacillus casei, Lactobacillus delbrueckii. Lactobacillus plantarum способен активно размножаться с хлебопекарными дрожжами, гомоферментативен, сбраживая сахара с образованием молочной кислоты. Оптимальная температура роста 30 °С, но может приспосабливаться и к пониженной температуре (15 °С). Развивается на средах, богатых витаминами и аминокислотами. Под микроскопом видны неподвижные палочки размерами (4...7)´ (0,5...0,7) мкм, группирующиеся парами или небольшими цепочками. При неблагоприятных условиях образуются длинные цепочки в виде изогнутых нитей. На плотных средах вырастают мелкие, гладкие, со слегка выпуклой сферической поверхностью грязно-белые колонии с зоной «просветления» при наличии мела. Бактерии спиртоустойчивы, растут при концентрациях спирта 15—20об.%. Lactobacillus fermenti активно размножается вместе с хлебопекарными дрожжами. Относится к гетероферментативным молочнокислым бактериям: продуцирует молочную, уксусную и муравьиную кислоты. Способен к слабому протеолизу белков. Требователен к витаминному составу питательных сред. Температурный оптимум 38—40 °С, плохо растет при 45 °С и не растет при пониженной температуре (15 °С). Бактерии имеют вид коротких палочек размером (2...4) ´ (0,5...1,0) мкм, группирующихся по две или небольшими цепочками. Lactobacillus brevis по основным признакам схож с Lactobacillus fermenti, но имеет более низкий температурный оптимум; специфичен для заквасок и активен в образовании ароматического комплекса хлеба. Lactobacillus casei относится к гомоферментативным бактериям и принимает активное участие в образовании молочной кислоты в бродящем тесте. По морфологическим, культуральным и физиологическим признакам близок к Lactobacillus plantarum. На рис. 2.22 приведена форма клеток бактерий Thermobacterium cereale (Дельбрюка), используемых при получении жидких дрожжей для подкисления питательной смеси и улучшения ее состава при размножении чистой культуры. Используют различные производственные расы бактерий, но все они имеют следующие морфологические, культуральные и физиологические признаки: палочки длиной 2,7—7 мкм и диаметром 0,4—0,7 мкм. В молодой культуре эти бактерии одиночные, иногда соединяются под прямым углом, изредка — в короткие цепочки, неподвижные, спор не образуют, грамположительные.

Lactobacillus delbrueckii (Э-1)имеет размер палочек (1...2,5) ´ 0,3 мкм. В зрелой культуре одиночные клетки соединяются в конгломераты от 8 до 20 и более палочек, расположенных в разных направлениях. Температурный оптимум кислотообразования 50 °С. Структура бактерий гораздо примитивнее, чем дрожжевых клеток. В бактериях функцию митохондрий выполняют особые образования цитоплазматической мембраны — мезосомы, специализирующиеся на выполнении различных функций. Некоторые из них участвуют в обмене липидов и углеводов, другие — в образовании клеточной оболочки. Часть мезосом является аналогом центросом и связана с аналогом ядра — нуклеотидом. Мезосомы богаты гидролитическими ферментами. Протекание жизненно важных процессов: переноса энергии и веществ, следовательно, связано с мембранами, которые состоят из белков и фосфолипидов. Рибосомы находятся в цитоплазме клеток на поверхности мембраны (тогда они активны) либо свободно плавают в цитоплазме; в состав рибосом входят рибонуклеопротеиды, т. е. РНК и белковый комплекс. В рибосомах содержится от 40 до 60 % белков и РНК. Белки рибосом имеют основный характер, в их составе преобладают лизин, аргинин и гистидин. Бактерии принадлежат к группе прокариотных микроорганизмов, у которых ядро не выражено, но имеется его аналог в виде нуклеотида или ядерного вещества, которое диффузно распределено в протоплазме. Источником энергии для бактерий является молочнокислое брожение, при котором из одной молекулы гексозы образуются две молекулы молочной кислоты:
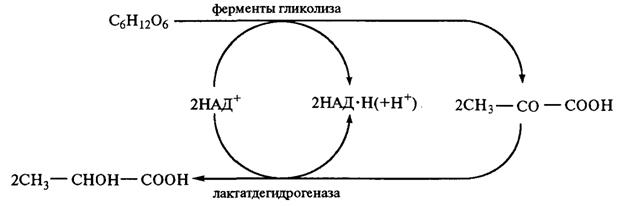
При молочнокислом брожении на каждую граммолекулу сброженной гексозы должна выделяться энергия, соответствующая 215,43 кДж теплоты (стандартное изменение свободной энергии). Сначала протекает гликолиз. В процессе анаэробного превращения гексозы в пируват расходуется две молекулы АТФ (в реакциях, катализируемых гексокиназой и фосфофруктокиназой). Вместе с тем на более поздних этапах процесса синтезируются четыре молекулы АТФ при превращении 1,3-дифосфоглицериновой кислоты в 3-фосфоглице-риновую и при превращении фосфофенолпирувата в енолпируват. Таким образом, количество энергии, запасаемой в виде АТФ при спиртовом и молочнокислом брожении, эквивалентно двум макроэргическим связям АТФ на 1 моль сброженной гексозы, что составляет 58 кДж. Затем пируват (СН3—СО—СООН) под влиянием фермента лактатдегидрогеназы (рис. 2.23) восстанавливается и образуется молочная кислота. Дегидрогеназы — ферменты, катализирующие реакцию дегидрирования, т. е. отщепление водорода от данного органического соединения. Лактатдегидрогеназа имеет кофермент НАД. Он является исключительно реакционноспособной окислительно-восстановительной системой, играющей важную роль в процессе спиртового и молочнокислого брожения. Вступая в соединение со специфическим белком, НАД образует анаэробную дегидрогеназу, обладающую способностью отнимать водород непосредственно от органических соединений. В результате происходит окисление данного соединения, при этом НАД превращается в свою восстановленную форму — НАД • Н2. Соединенный со специфическим белком НАД ■ Н2 обладает значительным восстановительным потенциалом. Он может передать свой водород уксусному альдегиду, образующемуся в качестве промежуточного продукта при спиртовом брожении или анаэробном дыхании. При этом ацетальдегид восстанавливается до этанола, а НАД • Н2 снова превращается в НАД. При молочнокислом брожении в процессе превращения углеводов ацетальдегид не образуется и НАД • Н2 осуществляет восстановление пирувата, который подвергается гидролизу пируват-карбоксилазой. В результате этой реакции, катализируемой лактатдегидрогеназой, пируват превращается в молочную кислоту и регенерируется НАД.
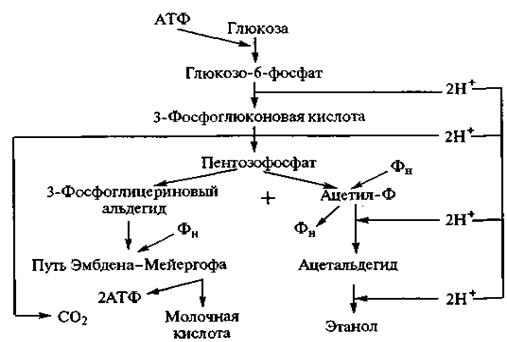
Рис. 2.23. Схема образования молочной кислоты Молочнокислые бактерии чаще всего встречаются в молочных продуктах. На их деятельности основано получение простокваши, сметаны и других кисломолочных продуктов. Они находятся и на зерне, поэтому ржаной хлеб можно получить после естественного, спонтанного размножения бактерий в тесте до обеспечения заданной кислотности. Это бактерии типа Escherichia aerogenes и Bacterium levans, близкие между собой виды. Развиваясь в среде, они образуют молочную кислоту, значительное количество летучих кислот и газов (водорода, азота и диоксида углерода). При производстве хлеба из ржаной муки размножение этих бактерий подавляется основной микрофлорой заквасок и высокой кислотностью среды. В промышленности молочную кислоту получают с помощью бактерий Bacterium delbriieckii (Lactobacillus delbrueckii), принадлежащих к термофильным. Оптимальная температура развития 45—50 °С. В микроскопе они видны в виде длинных палочек. Молочнокислое брожение играет очень большую роль при производстве молочнокислых продуктов, изготовлении кваса, хлебных заквасок и жидких дрожжей, в технологии хлеба, при квашении овощей. Все микроорганизмы, вызывающие молочнокислое брожение, разделяются на две большие группы. К первой группе принадлежат бактерии, подобные Streptococcus lactis, Streptobacterium plantarum, и палочки типа Thermobacterium cereale (Bacterium delbrueckii), являющиеся настоящими анаэробами и сбраживающие гексозы в соответствии с вышеприведенным уравнением молочнокислого брожения. Эти молочнокислые бактерии называют гомоферментативными. Гомоферментативное молочнокислое брожение протекает по гликолитической схеме. В отличие от спиртового брожения на последнем этапе гликолиза под действием лактатдегидрогеназы пируват восстанавливается до лактата. Streptobacterium plantarum — короткие, неподвижные, неспособные к образованию спор палочки длиной 2—3 мкм и диаметром 1 мкм, образуют короткие цепочки. Благоприятная температура развития 30—35 °С. Молочнокислые бактерии, являясь живыми организмами, имеют комплекс ферментов, которые могут повлиять на характеристики хлебопекарного теста, осуществляя протеолиз и даже амилолиз. К гомоферментативным молочнокислым бактериям относятся Lactobacillus casei и Lactobacillus plantarum. Гомоферментативные молочнокислые бактерии принимают активное участие в образовании молочной кислоты в бродящем тесте. Ко второй группе бактерий принадлежат микроорганизмы, которые кроме молочной образуют яблочную, пировиноградную, янтарную, уксусную и другие кислоты, этанол, диоксид углерода и др. К ним относятся Lactobacillus brevis и Lactobacillus fermenti — гетероферментативные молочнокислые бактерии. В качестве источников энергии эти бактерии используют глюкозу, фруктозу и мальтозу. На образование янтарной и уксусной кислот, этанола и диоксида углерода бактерии затрачивают до 50 % сбраживаемых сахаров. В качестве источника энергии они могут использовать и органические кислоты — лимонную, яблочную и даже молочную. Промежуточными продуктами метаболизма гетероферментативных молочнокислых бактерий являются спирты, эфиры, карбонильные соединения, выход которых у некоторых штаммов достигает нескольких процентов. В формировании аромата хлеба эти вещества играют определенную роль. Гетероферментативные бактерии характеризуются принципиально другим, чем гомоферментативные, путем разложения глюкозы — пентозофосфатным. По Буду, схема процесса гетероферментативного молочнокислого брожения, осуществляемая Leuconostoc mesenteroides и Lactobacillus brevis, имеет следующий вид:
Более детальное исследование ферментов окислительного фосфорилирования у ряда штаммов Leuconostoc mesenteroides и Lactobacillus brevis в бесклеточных экстрактах показало, что декарбоксилирование 6-фосфоглюконата может идти путем прямого декарбоксилирования субстрата с образованием рибулозо-5-фосфата, а у некоторых штаммов — путем дегидрирования 6-фосфоглюконата в 2-кето-6-фосфоглкжонат с последующим де-карбоксилированием последнего. Питательные смеси, в которых осуществляется культивирование микроорганизмов, оказывают существенное влияние на их жизнедеятельность, индукцию ферментов, качество жидких дрожжей и жидких заквасок, биотехнологические показатели этих полуфабрикатов. Принципиальной особенностью кинетики микробных популяций является зависимость скорости роста микроорганизмов от концентрации наиболее важных компонентов среды, обеспечивающих биосинтетическую основу метаболизма. Скорость изменения числа микроорганизмов в режиме роста (в экспоненциальной фазе) линейно связана с концентрацией клеток в системе: dN/dt = mN, (2.1) где N — число клеток; t — продолжительность процесса; m — коэффициент пропорциональности, или удельная скорость роста, m= (1/N)dN/dt, имеет размерность, обратную времени. Предполагается, что m не зависит от времени в исследуемом интервале. Интегрирование уравнения (2.1) при начальном условии t = 0, N= N0 приводит к функции N= N0emt. (2.2) Удельная скорость роста (m) зависит от концентрации лимитирующего субстрата S: mS =mmaxS/(Ks+S), (2.3) где mmax — предельная максимальная удельная скорость роста; Ks — константа сродства субстрата к микроорганизму. При «насыщающих» концентрациях субстрата максимальная скорость роста mmax = аR/(аR +а)1п2(kкатЕ/В), (2.4) где aR — коэффициент пропорциональности, равный константе скорости второго порядка; kкат — каталитическая константа скорости; E — ферментная система (фермент); В — содержание базовой ДНК, моль оснований ДНК на 1 клетку.
Уравнение (2.4) свидетельствует о том, что основным фактором, приводящим к существенному уменьшению mmax по сравнению с kкат,является распределение потоков ключевого метаболизма [аR /(aR + а) может быть существенно меньше единицы, если a/aR>>1], т.е. если на репликационный процесс идет небольшая доля вещества, а в клетке содержится небольшое количество лимитирующего фермента по сравнению с ДНК. На кинетику роста популяции оказывают влияние транспортные процессы. Перемещение лимитирующего субстрата обеспечивается под действием пермеаз. Характерным представителем группы гетероферментативных бактерий являются Bacterium lactis aerogenes, образующие молочную, уксусную кислоты, этанол, диоксид углерода, водород и метан. Выход уксусной кислоты при сбраживании Сахаров подобными микроорганизмами может превышать выход молочной кислоты. Из группы гетероферментативных бактерий в заквасках и в тесте постоянно размножаются β-бактерии. Это палочки разной длины: от очень коротких — длиной 1,1—1,5 мкм и диаметром 1 мкм до длинных — соответственно 2,5—15 и 1,2—2 мкм, неподвижные, спор не образуют. Продуктами их метаболизма кроме молочной кислоты являются уксусная кислота и иногда довольно значительное количество диоксида углерода. Оптимальная температура роста 25 °С. Эти бактерии участвуют в созревании ржаного теста, обеспечивая не только накопление кислот в нем, но и его разрыхление. В последние годы в производстве ржаного и ржано-пшеничного хлеба применяют закваски, приготовленные на чистых культурах молочнокислых бактерий: Lactobacillus plantarum, Lactobacillus brevis, Lactobacillus casei, Lactobacillus fermenti. Заметное содержание молочной и уксусной кислот в ржаном тесте и хлебе объясняется тем, что при брожении ржаного теста основным является молочнокислое, при котором накапливается как молочная, так и уксусная кислота. Особенно большое количество молочной кислоты образуется при сбраживании сахара некоторыми термофильными молочнокислыми микробами, подобными широко применяемым в пищевой промышленности Thermobacterium cereale (Bacterium delbriieckii). Определенное значение имеет тот факт, что одни молочнокислые бактерии образуют оптически недеятельную молочную кислоту, другие — D-форму и третьи — L-форму. При этом следует отметить, что один и тот же микроб при культивировании его на разных питательных средах образует различные формы молочной кислоты. Так, Lactobacillus casei при развитии на моносахаридах и лактозе образует правовращающую молочную кислоту, а на сахарозе и мальтозе — оптически недеятельную левовращающую форму. Некоторые микроорганизмы не могут строить белки из неорганических источников азота, и для роста и развития им требуется целый ряд аминокислот, которые усваиваются ими в неизменном виде, называемых незаменимыми. К числу таких микроорганизмов относятся и молочнокислые бактерии. В зависимости от физиологических особенностей микробов количество незаменимых для них аминокислот различно. Так, например, для золотистого стафилококка обязательно наличие в питательной среде двух аминокислот: триптофана и цистина; для молочнокислых бактерий Lactobacillus casei обязательно наличие 16 аминокислот: лизина, аргинина, гистидина, фенилаланина, тирозина, триптофана, аланина, валина, лейцина, изолейцина, серина, треонина, цистина, метионина, аспарагиновой и глутаминовой кислот. Аминокислоты, ассимилируемые микроорганизмами в неизменном виде, играют двойную роль: как вещества, необходимые для построения белков, содержащихся в теле данного микроба, или же как вещества, необходимые для построения активных групп ферментов. В последнем случае аминокислоты являются своего рода витаминами. Так, например, глутаминовая кислота участвует в построении ферментов, катализирующих синтез пу-риновых оснований и пептидов у молочнокислых бактерий Lactobacillus casei. Некоторые молочнокислые бактерии весьма интенсивно де-карбоксилируют глутаминовую кислоту с образованием диоксида углерода и g-аминомасляной кислоты:
Следовательно, в результате действия декарбоксилазы на глутаминовую кислоту образуется новая аминокислота. Декарбоксилирование аминокислот играет, по-видимому, важную роль в обмене веществ микроорганизмов. Активная группа в декарбоксилазах аминокислот у бактерий представляет собой пиридоксальфосфат. Таким образом, важный этап диссимиляции аминокислот, каким является их декарбоксилирование, осуществляется в молочнокислых бактериях за счет активных групп соответствующих ферментов. Молочнокислые бактерии, развиваясь в природных и производственных субстратах, вступают в сложное взаимодействие с другими микроорганизмами. При этом существенную роль играет их видовая принадлежность. Заметное содержание молочной и уксусной кислот в ржаных полуфабрикатах хлеба объясняется тем, что при создании этих полуфабрикатов в них одновременно происходит как спиртовое, так и молочнокислое брожение. Это объясняется присутствием в заквасках и тесте как дрожжевых клеток, вызывающих спиртовое брожение, так и бактерий, вызывающих молочнокислое брожение. Подобного рода совместное существование этих микроорганизмов (симбиоз), оказывающих друг на друга положительное влияние, наблюдается при производстве целого ряда полуфабрикатов из них — в хлебном квасе, кумысе, жидких хлебопекарных дрожжах, жидких и густых ржаных заквасках, различных молочнокислых продуктах — айране, кавказском мацони, кефире. |
||||
|
|
Последнее изменение этой страницы: 2018-04-12; просмотров: 1138. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |