|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
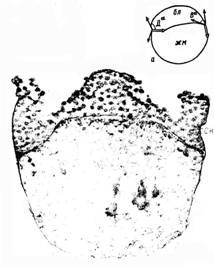
Морфологические маркеры дорсовентральной полярности, основанные на разнице механических напряжений; итоги операций (парафронтальные надрезы) на поздней высокой бластуле. (Сивкова)Долгое время всех интересовал вопрос, как же отличить дорсальную и вентральную стороны в зародыше рыб. В итоге выяснили, что есть некоторые маркёры: если посмотреть на зародыш на стадии высокой бластулы, то можно заметить некоторые различия в форме бластодермы на Д и В сторонах. Если провести две касательные (к бластодерма и к поверхности ЖК), выходящие из границы между бластодермой и ЖК, то можно заметить, что на Д стороне угол между этими касательными более острый, чем на В стороне, и образуется как бы «шапка с козырьком». Напомню, что в зародыше есть механические напряжения, они разные на разных стадиях, и они могут выступать как причины дальнейших дифференцировок клеток. Так вот, на Д стороне механические напряжения намного сильнее. Если нарушить целостность бластодермы (сделать надрез), то участок зародыша отгибается наружу, как бы сбрасывает напряжение. По степени отгиба участков можно сказать, Д это сторона или вентральная: на Д стороне степень отгиба намного выше, а на В почти и не отгибается, участок как будто бы продолжает окружность ЖК (опыты четы Черданцевых). Так же можно поиграть с напряжениями, что приведёт в переориентации осей. Например, если удалить немного бластодермы с В части зародыша, то клетки все направятся в тут сторону, чтобы поскорее закрыть рану, напряжение на В стороне резко повысится и впоследствии щиток может образоваться именно там.
Раздел «Производные мезодермы»
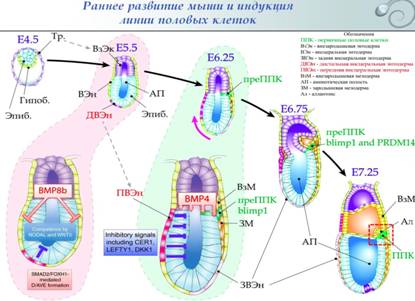
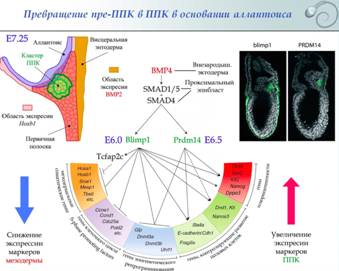
1. Формирование и миграция первичных половых клеток у млекопитающих. Даша В эмбриогенезе млекопитающих ППК возникают на границе эпибласта, первичной полоски и аллантоиса под индукционным влиянием лежащей проксимальнее («надлежащей») внезародышевой эктодермы. Индукционные процессы (секреция BMP8 внезародышевой эктодермой) начинаются уже на 5,5 сутки после оплодотворения. Эти сигналы являются подготовительными и не приводят непосредственно к образованию ППК. На 6-е сутки внезародышевая эктодерма осуществляет секрецию BMP4. При этом воздействие оказывается только на постериорную сторону зародыша: клетки проксимальной внезародышевой энтодермы секретируют агенты, ингибирующие BMP4 и индуцирующие экспрессию CER1, LEFTY1, DKK1 в антериорном эпибласте. Таким образом, возникает антериопостериорный градиент BMP4. На постериорной стороне, где BMP4 больше, на границе внезародышевой эктодермы и эпибласта возникают предшественник ППК.  BMP4 через посредников SMAD1/5 и SMAD4 индуцирует экспрессию Blimp1 на 6-е сутки и Prdm14 на 6,5 сутки. Продукты этих генов являются транскрипционными факторами и изменяют профиль экспрессии генов в преППК. Blimp1 и Prdm14 обладают одинаковыми эффектами ● Происходит снижение уровня экспрессии мезодермальных соматических генов (Hoxa1, Hoxb1 (экспрессируется по всей первичной полоске (постериорному эпибласт-у), и выключается только в кластере преППК), Snail1, Tbx6, etc.), генов клеточного цикла (S-phase promoting factors: Cdc25a, Ccnd1, etc.) и генов эпигенетического репрограммирования (Glp, Uhrf1, etc.); ● Повышается уровень экспрессии генов плюрипотентности (Oct4, Sox2, Klf2, Nanog, Dppa3) и генов, регулирующих развитие половых клеток (Kit –рецепторная тирозин-киназа , блокирует апоптоз и поддерживает регенерацию; лиганд к Kit – Stella, важен для миграции: при нокауте stella ППК гибнут при миграции).
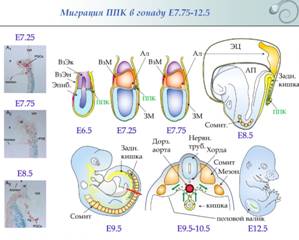
Миграция осуществляется по задней кишке к половому валику, который формируется как утолщение целомического эпителия на мезонефросе.
В течение первых 9 дней ППК мигрируют из постериорной части эпибласта через эндодерму постериорного участка первичной полоски в заднюю кишку. На 9й день они покидают заднюю кишку и на следующий день осуществляют миграцию к половым валикам, которые заселяют на 11,5 день. Как и мигрирующие ППК Xenopus, ППК млекопитающий тесно взаимодействуют с клетками, по которым они мигрируют, и миграцию они осуществляют, распространяя филаподии по субстрату. Направляющие миграцию механизм неясен. Предполагается, что основным субстратом миграции является фибронектин и, соответственно, экспрессируемые мигрирующими ППК Е-кадгерины. Вместе с ППК мигрируют SCF (stem cell factor)-секретирующие клетки, формируя, таким образом, «мигрирующую нишу». SCF играет важную роль в поддержании жизнеспособности и мобильности ППК. ППК способны к трансмиграции через одноклеточные тканевые пласты.
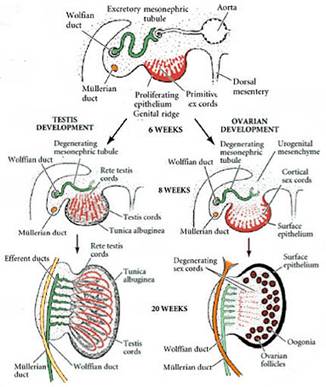
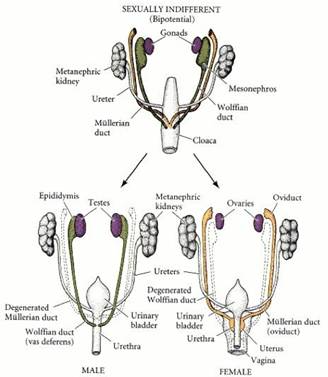
2. Первичная детерминация пола у млекопитающих. Даша Это детерминация гонад и образование яичника и семенника из бипотенциальной гонады. Чисто хромосомная, как правило, не зависит от внешнего воздействия. Первичная детерминация не происходит “по умолчанию”, формирование яичников и семенников - активные, контролируемые генами процессы. Вот в отсутствие гонад у млекопитающих (нет первичной генетической детерминации) формируется фенотип самки. Y-хромосома несет ген тестис-детерминирующего фактора образования семенников. Гонадогенез (бипотентный гонадный валик): 9.5-15 суток у мыши, 4-я неделя - у человека; закладка в виде парных гребневидных утолщений участков промежуточной мезодермы; до 7-й недели у человека зачатки гонад индифферентны по половым признакам. В течение индифферентной стадии (бипотентной) эпителий половых валиков пролиферирует и образует первичные половые тяжив мезенхиме валика. ППК заселяют гонады на 6-й неделе (~35-й день) у человека, 7.75-12.5 суток у мыши. Если генотип плода XY - тяжи углубляются внутрь зачатка гонады и образуют половые тяжи семенника, которые впоследствии утрачивают контакт с поверхностным эпителием и отделяются от него слоем ВКМ - tunica albuginea - белочной оболочкой. Т.о. когда ППК попадают в мужские гонады, они будут далее развиваться в тяжах внутри семенника. У ХХ-гонад первичные половые тяжи дегенерируют, но эпителий производит кортикальные половые тяжи, которые не проникают в мезенхиму и остаются вблизи наружной поверхности гонады. ППК станет яйцом, а окружающий ее кластер кортикальных половых тяжей - клетками гранулезы. Мезенхимные клетки яичника дифференцируются в клетки теки. Вместе клетки теки и гранулезы с яйцом образуют фолликулы. У XX-плодов сохраняется Мюллеров проток, дифференцируется в яйцеводы, матку, шейку матки и верхнюю часть влагалища. Вольфов проток в отсутствие тестостерона дегенерирует. У XY-плодов Вольфов проток дает структуры эпидидимиса и семявыносящего протока, протока семенного пузырька. Мюллеров дегенерирует.
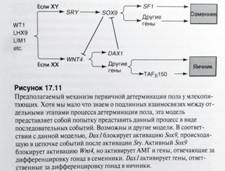
Механизмы и факторы первичной детерминации. Покане очень ясно, но накоплены следующие данные:
● факторSRY(ген sex-determining region of the Y chromosome) у человека и его гомолог Sry у мышей - главный по формированию семенников; есть только у млекопитающих; ген расположен на коротком плече Y-хромосомы. Особи, рожденные с Y-хромосомой только с длинным плечом, - фенотипически женского пола. Белок SRY у человека обнаружен в сплайсосомах и участвует в процессировании пре-мРНК, также в клетках Сертоли. Но показано, что SRY/Sry ● необходим, но недостаточен для формирования полноценных семенников; ● Sox9- аутосомный ген, детерминирующий семенники; есть у всех позвоночных; возможный механизм действия - как фактор сплайсинга и транскрипционный фактор; содержит ДНК-связывающий HMG-домен (high mobility group box). Лица с генотипом ХХ, несущие дополнительные копии Sox9, развиваются по мужскому типу (хотя не имеют ни одной копии гена SRY!). Т.о. Sox9 может функционально замещать SRY в детерминации семенников. Это неудивительно, все-таки Sox9 есть у всех позвоночных - более древний ген. У млеков он активируется с помощью Sry, и возможный механизм - Sry “включатель”, а белок Sox9 инициирует эволюционно консервативный путь образования семенников; ● FGF9- может быть одной из мишеней воздействия генов Sry или Sox9; сам FGF9 индуцирует миграцию мезонефротических клеток в гонаду, важен для стимуляции деления клеток Лейдига (тестостерон!), ускорения дифференцировки клеток Сертоли. У нокаутных по гену Fgf9 мышей почти все гомозиготы являются самками; ● SF1(стероидогенный фактор 1) -транскрипционный фактор, необходим для образования бипотентной гонады, его активность сохраняется в формирующихся семенниках. Участвует в повышении уровня транскрипции гена АМГ (Антимюллеров гормон) в клетках Сертоли, в стимуляции продукции тестостерона в клетках Лейдига; ● DAX1 - ген Х-хромосомы, способный угнетать образование семенников. Открыт и описан у двух XY сестер. Первоначально экспрессируется в гонадных валиках у мышей обоего пола, далее только в зачаточной ХХ-гонаде. Противодействует Sry и Sox9, снижает уровень экспрессии SF1. ● WNT4 - аутосомный ген, способный детерминировать образование яичников. Экспрессируется в бипотентных гонадных валиках, далее сохраняется в ХХ-гонадах; по-видимому, его продукт активирует ген Dax1. Возможная модель детерминации пола:
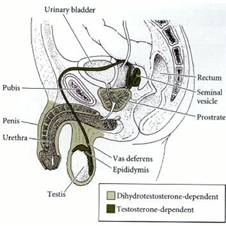
3. Вторичная детерминация пола у млекопитающих. Гормональная регуляция полового фенотипа. Даша Определяет фенотип особи вне гонад, охватывает развитие фенотипа самки/самца в ответ на действие гормонов половых желез. Затрагивает системы выводящих протоков и канальцев самцов и самок, формы гениталий, половые различия по размерам, голосовым связкам (типам хряща), по мускулатуре. Вторичные половые признаки обычно контролируются гормонами половых желез. Выделяют 2 периода вторичной детерминации: в ходе органогенеза зародыша и в период полового созревания. Для образования мужского фенотипа необходимо 2 гормона: ● АМГ (антимюллеров гормон) - клеток Сертоли; член семейства TGF-β, вызывает разрушение Мюллерова протока, участвует в индукции Вольфова протока к дифференцировке в эпидидимис, vas deferens, семенные пузырьки, стимулирует развитие мочеполовых валиков в мошонку и пенис; ● тестостерон - клетки Лейдига; отвечает за дифференцировку Вольфова протока (+угнетает развитие молочных желез); превращается в 5α-дигидротестостерон в мочеполовом синусе и семенных пузырьках - более сильный гормон по сравнению с тестостероном, отвечает за формирование наружных половых органов. Если по каким-либо причинам отсутствует функционально активный ген фермента, превращающего тестостерон в 5α-дигидротестостерон, вспоминайте историю Аси об общине в Доминиканской республике: дети ХY с семенниками (и вообще мужскими внутренними анатомическими признаками), но со слепым вагинальным карманом, увеличенным клитором и с девичьей внешностью. ● Insl3 - инсулиноподобный гормон 3 клеток Лейдига; требуется для опускания гонад в мошонку; самцы, не имеющие этого гормона, бесплодны. У самок отсутствие этого гормона приводит к нарушениям менструального цикла. Районы мужской половой системы, развитие которых зависит от действия тестостерона и дигидротестостерона: У людей XY с синдромом андрогенной нечувствительности (не реагируют на гормоны маскулинизации, нет рецепторов) формируется женский фенотип, поскольку организм подвергается воздействию эстрогена. Но анатомически - есть семенники и, следовательно, “бесполезный” для них тестостерон; если сохраняется чувствительность к АГМ, то Мюллеровы протоки дегенерируют, как и положено. Это явление псевдогермафродитизма - гонады одного типа, а вторичные половые признаки - другого. Такие лица развиваются как фенотипически нормальные, но стерильные женщины, без матки и яйцеводов, с семенниками в брюшной полости. “Истинные” гермафродиты содержат ткани мужской и женской гонад. Про самок. Эстроген - секретируется фолликулярными клеткамми яичника.Необходим для полноценного формирования как мюллерова, так и вольфова протоков. Просто у самок эстрогена, секретируемого фетальными яичниками, больше и достаточно для индукции дифференцировки Мюллерова протока на матку, шейку матки и яичники. Эстроген необходим для формирования фертильных самцов и самок. У нокаутных по генам рецепторов эстрогена мышей: у самок - ПК погибают при достижении половозрелости, а клетки гранулезы начинают развиваться как клетки Сертоли; у самцов - образуется очень мало спермы (эстроген у них регулирует абсорбцию воды в семявыносящем протоке для концентрирования и сохранения спермы). Эстроген ускоряет развитие молочных желез. Несмотря на то, что концентрация эстрогена в крови самок выше, чем в крови самцов, концентрация эстрогена в сети семенников даже выше, чем в крови самок!
4. Бипотенциальная (индифферентная) стадия развития. Даша Гонадогенез (бипотентный гонадный валик): 9.5-15 суток у мыши, 4-я неделя - у человека; закладка в виде парных гребневидных утолщений участков промежуточной мезодермы; до 7-й недели у человека зачатки гонад индифферентны по половым признакам. Из бипотенциальной гонады в результате первичной детерминации пола формируются либо яичники, либо семенники. В течение индифферентной стадии (бипотентной) эпителий половых валиков пролиферирует и образует первичные половые тяжив мезенхиме валика. ППК заселяют гонады на 6-й неделе (~35-й день) у человека, 7.75-12.5 суток у мыши. Если генотип плода XY - тяжи углубляются внутрь зачата гонады и образуют половые тяжи семенника, которые в последствие утрачивают контакт с поверхностным эпителием и отделяются от него слоем ВКМ - tunica albuginea - белочной оболочкой. Т.о. когда ППК попадают в мужские гонады, они будут далее развиваться в тяжах внутри семенника. У ХХ-гонад первичные половые тяжи дегенерируют, но эпителий производит кортикальные половые тяжи, которые не проникают в мезенхиму и остаются вблизи наружной поверхности гонады. ППК станет яйцом, а окружающий ее кластер кортикальных половых тяжей - клетками гранулезы. Мезенхимные клетки яичника дифференцируются в клетки теки. Вместе клетки теки и гранулезы с яйцом образуют фолликулы. У XX-плодов сохраняется Мюллеров проток, дифференцируется в яйцеводы, матку, шейку матки и верхнюю часть влагалища. Небольшие верхние участки Мюллерова протока сохраняются и у самцов и образуют morgagni hydatida (он же appendix testis;срастается с семенником) и vagina masculina (мужскя маточка; открывается в простату). Вольфов проток в отсутствие тестостерона дегенерирует. У XY-плодов Вольфов проток дает структуры эпидидимиса и семявыносящего протока, протока семенного пузырька. Мюллеров дегенерирует.
5. SRY – главный половой фактор Y-хромосомы. Даша ФакторSRY(ген sex-determining region of the Y chromosome) у человека и его гомолог Sry у мышей - главный по формированию семенников; есть только у млекопитающих; ген расположен близко к концу короткого плеча Y-хромосомы. Особи, рожденные с Y-хромосомой, представленной только коротким плечом - мужского пола, а только с длинным плечом - фенотипически женского пола. Этот ген обнаружен у нормальных мужчин ХУ и редко встречающихся с генотипом ХХ; отсутствует у нормальных женщин ХХ и у многих женщин с аномальным генотипом ХУ. Остальная часть женщин с генотипом ХУ несет точечные мутации или мутации типа “сдвиг рамки считывания” гена SRY, эти мутации не позволяют белку SRY связываться с ДНК и работать на образование семенников. Сам пептид, кодируемый SRY, по-видимому, транскрипционный фактор, т.к. содержит ДНК-связывающий домен HMG-бокс (high mobility group box). Белок SRY у человека обнаружен в сплайсосомах и участвует в процессировании пре-мРНК, также в клетках Сертоли. Мыши ХХ, трансгенные по гену Sry, являются самцами. Показано, что SRY/Sry необходим, но недостаточен для формирования полноценных семенников. Существует ряд аутосомных генов, участвующих в детерминации судьбы полового зачатка (см. вопрос 2).
|
||
|
|
Последнее изменение этой страницы: 2018-04-12; просмотров: 479. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |