|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
Особенности окисления ненасыщенных жиных кислотДо двойных связей окисление ненасыщенных жирных кислот происходит так же, как окисление насыщенных жк.
Синтез жирных кислот Имеет ряд особенностей: 1. синтез ЖК в организме человека начинается с пальмитиновой к-ты (С16), затем происходят реакции удлинения углеродного скелета и дегидрирования. 2. в организме человека не синтезируются ЖК с двойными связями, расположенными дистальнее С9, поэтому их необходимо получать с пищей (эссенциальные ЖК). 3. в отличии от окисления синтез ЖК локализован в эндоплазматической сети. 4. источником синтеза является малонил-КоА, который образуется из ацетил-КоА 5. для восстановления промежуточных продуктов синтеза ЖК используются НАДН2 (ПФЦ). 6. все реакции биосинтеза ЖК катализируются полиферментным комплексом (пальмитат синтазой), локализованным в цитозоле клетки и реакции протекают на поверхности этого комплекса.
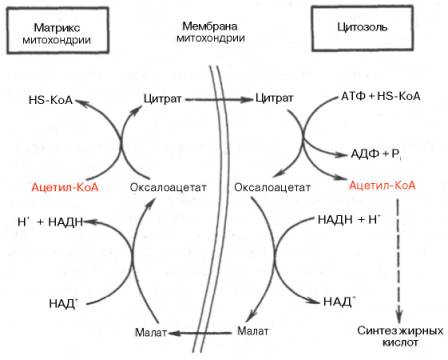
СТАДИЯ Образование малонил-КоА для синтеза ЖК Его субстратом является ацетил-КоА, образующийся в результате окислительного декарбоксилирования ПВК.  Мембрана митохондрий непроницаема для Ацетил-КоА. Транспорт Ацетил-КоА в цитоплазму может происходить: а) с помощью карнитина (аналогично переносу ЖК) б) Ацетил-КоА в митохондриях преобразуется в цитрат (ацетил-КоА+ЩУК), затем цитрат проходит в цитоплазму, там опять распадается на ЩУК и ацетил-КоА из которого образуется малонил-КоА, Е ацетил-КоА-карбоксилаза, КоЕ биотин, Мg²
СТАДИЯ Синтеза ЖК на поверхности пальмитатсинтетазы
Пальмитатсинтетаза состоит из 7 ферментов, в центре ацилпереносящий белок (АПБ), по переферии 6 ферментов. Комплекс имеет две свободные SН-группы, в том числе одну на АПБ. Обе SН-группы являются акцепторами ацилов. Обозначим пальмитатсинтетазу
1. Реакция трансацилирования Е-комплекса
2. Перенос малонила с малонил КоА на синтетазу
3. Р-ция конденсации ацетила с малонилом и декарбоксилирование образовавшегося продукта, Е β-кетоацилсинтетаза
4. 1-ое восстановление промежуточного продукта с участием НАДФН2
Е β-кетоацилредуктаза
5. Дегидратация промежуточного продукта
Е гидроксиацилгидратаза
6. 2-ое восстановление промежуточного продукта
Е еноилредуктаза
7. Синтезированный бутирил с помощью 1-го фермента ацетилтрансацилазы переносится на свободную SН-группу (верхнюю). На нижнюю SН-группу поступает новый малонильный остаток. Цикл повторяется. Для синтеза пальмитиновой кислоты требуется 7 таких циклов, т.е. 7 остатков малонила и 1 ацетила-КоА. Суммарное уравнение биосинтеза пальмитиновой кислоты. 1 ацетил-КоА+7малонил-КоА+14НАДФН2 → пальмитиновая к-та+8НS-КоА+14НАДФ+6Н2О+7СО2
Гидролиз ферментативного комплекса с синтезируемой ЖК происходит под действием пальмитатдеацилазы. Удлинение ЖК происходит путем добавления ацетильных остатков в митохондриях и путем добавления малонильных остатков в цитоплазме.
ТЕМА: ЛИПИДЫ III Цель:Дать представление о механизмах регуляции липидного обмена, патологии, возникающие в результате его нарушения.
ПЛАН ЛЕКЦИИ 1. Регуляция обмена липидов. Регуляция окисления и синтеза ЖК. 2. Метаболизм кетоновых тел в норме и патологии 3. Транспортные липопротеины (ЛП) 4. Липопротеинемии и атеросклероз. 5. Депонирование и мобилизация жиров. 6. Распространение и функции Хс. 7. Синтез Хс. 8. Транспорт Хс. 9. Сложные липиды и миелинизация.
Регуляция обмена липидов Интенсивность обмена липидов в тканях зависит от: а) поступления липидов с пищей; б) нервно-гумморальной регуляции Избыточное поступление углеводов и ТАГ с пищей препятствует расходу эндогенных запасов ТАГ в жировой ткани. Прием даже полностью углеводной пищи существенно влияет на синтез ТАГ и Хс. Вместе с тем, растительные масла, содержащие ненасыщенные ЖК напротив, способствуют мобилизации липидов. Наличие липотропных факторов в пище облегчает биосинтез фосфолипидов, а их отсутствие способствует образованию ТАГ. Мобилизации ТАГ из жирового депо и угнетению биосинтеза холестерина способствуют голодание и эмоциональный стресс.
Нервно-гормональная регуляция липидного обмена Стимуляторы тканевойлипазы - адреналин, норадреналин, глюкагон, тироксин, адренокортикотропный гормон(активируют аденилатциклазу) Ингибитор липолиза – инсулин (угнетает аденилатциклазу). В результате воздействия гормонов нервной клетки рецептор модифицирует свою структуру. В такой форме данный рецептор способен активировать аденилатциклазу, что стимулирует образование ц-АМФ и АТФ. Далее ц-АМФ активирует протеинкиназу, а последняя, путем фосфорилирования неактивную тканевуюлипазу превращает в активную. |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2018-05-10; просмотров: 409. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||