|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
Компоненты нуклеиновых кислот
Структура пиримидиновых и пуриновых оснований
Пиримидиновые и пуриновые азотистые основания, входящие в состав нуклеотидов, представляют собой замещенные производные шестичленного гетероцикла – пиримидина, с одной стороны, и пурина –сложной гетероциклической системы, состоящей из двух конденсированных гетероциклов: пиримидина и имидазола, с другой стороны (Рис. 12.1).
Рисунок 12.1 – Структурные формулы пиримидина и пурина
Положения атомов в ароматических кольцах имеют нумерацию в соответствии с номенклатурой, принятой Международным союзом теоретической и прикладной химии (International Union of Pure and Applied Chemistry – IUPAC) и Международным союзом биохимии и молекулярной биологии (International Union of Biochemistry and Molecular Biology – IUBMB). Следует обратить внимание на то, что нумерация в пиримидиновом и пуриновом кольцах идет в противоположных направлениях, при этом атом углерода С5 в обеих молекулах находится в одном и том же положении.
Азотистые основания нуклеиновых кислот
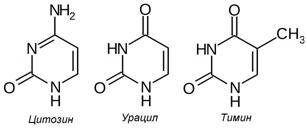
Пиримидиновые основания как у прокариот, так и у эукариот представлены цитозином (Cyt), урацилом (Ura) и тимином (Thy), структурные формулы которых приведены на Рис. 12.2. Пуриновые основания представлены аденином (Ade) и гуанином (Gua). ДНК содержит два пиримидиновых основания – цитозин и тимин и два пуриновых основания – аденин и гуанин. В состав РНК также входят два пурина – аденин и гуанин и два пиримидина – цитозин и урацил.
Рисунок 12.2 – Структурные формулы пуриновых и пиримидиновых оснований нуклеиновых кислот 
Кроме аденина и гуанина известны два других пуриновых основания – гипоксантин (Hyp) и ксантин (Xan), которые выступают в роли интермедиатов в процессах метаболизма пуринов, в частности гипоксантин и ксантин являются продуктами окислительного дезаминирования аденина и гуанина, соответственно. Гипоксантин выполняет важную функцию в качестве одного из оснований входящих в состав антикодонов ряда транспортных РНК. У человека в роли конечного продукта катаболизма пуринов выступает окисленное пуриновое основание – мочевая кислота (UA).
Рисунок 12.3 – Структурные формулы минорных пуриновых оснований
Физико-химические свойства пуриновых и пиримидиновых оснований
Явление таутомерии
Благодаря феномену кето-енольной таутомерии азотистые основания в нуклеотидах могут существовать либо в лактимной, либо в лактамной формах. В физиологических условиях преобладает лактамная форма гуанина тимина и урацила, но лактимные формы аденина и цитозина. Для осуществления специфического спаривания основания должны находиться в соответствующей таутомерной форме. Миграция водородного атома позволяет каждому основанию существовать в различных таутомерных формах. Основания, в составе двойной спирали ДНК, образующие канонические пары, должны иметь амино-группы (-NH2) и кето-группы (>С=О) в отличие от таутомеров, имеющих имино-группы (=NH) и енольные группы (-ОН) и способных к неканоническому спариванию, как, например, пуриновое основание Ade, который может образовать пару с таутомерной имино-формой Cyt (Рис. 12.4).
Рисунок 12.4 – Образование неканонических пар с участием аденина и имино-формы цитозина (А) и нормального цитозина с имино-формой аденина (Б)
Как известно, точность копирования в процессе репликации ДНК настолько велика, что в среднем на каждые 1·109 пар нуклеотидов приходится всего одна ошибка. Такую высокую точность репликации обеспечивает корректирующая (3¢®5¢)-экзонуклеазная активность ДНК-полимеразы. Тем не менее, в ряде случаев ДНК-полимераза может ошибаться. Как раз одной из причин таких ошибок является способность всех азотистых оснований образовывать термодинамически невыгодные таутомерные формы за счет миграции атома водорода. При этом как указывалось выше, амино- и оксогруппы превращаются в имино- и енольные группы, соответственно. Такие редкие таутомерные формы, как правило, образуют неправильные, неканонические пары с другими основаниями. Примеры такого спаривания показаны на Рис. 12.4. Так, имино-форма Cyt образует пару не с Gua, а с Ade. В результате, в процессе последующей репликации, может произойти замена пары A-T на G-C. Точно также Ade способен образовывать редкую таутомерную имино-форму, которая приобретает способность комплементарно спариваться с неканоническим для него Cyt. В норме образование пар между двумя пуринами, двумя пиримидинами или некомплементарными основаниями A-C или G-T стерически затруднено, поскольку при этом не могут образовываться подходящие водородные связи и, следовательно, нарушается геометрия спирали. Модифицированные пурины и пиримидины, с небольшой частотой встречающиеся в ДНК, образуют такие же водородные связи, что и их немодифицированные аналоги. В этом случае правила спаривания не нарушаются.
|
|||||||||||
|
|
Последнее изменение этой страницы: 2018-04-12; просмотров: 524. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |