|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
Внутриклеточный синтез липидовСтр 1 из 6Следующая ⇒ Гидролиз и всасывание белков
Взрослые потребляют с пищей 70-90 г белков в день, а детям в расчете на 1 кг веса их требуется в 5-10 раз больше. Почти такое же количество белков поступает в просвет кишечника в составе пищеварительных соков, слущивающихся клеток и в виде белков сыворотки. При синдроме экссудативной энтеропатии через кишечник теряется так много белка плазмы, что его убыль не компенсируется синтезом новых белков в печени, и возникает гипопротеинемия. Ферментативный гидролиз. Переваривание белков начинается в желудке, но роль этого этапа относительно невелика, посколькупепсином гидролизуется не более 10-15% белков пищи. У больных, страдающих ахилией и дефицитом пепсина, белки, тем не менее, могут нормально перевариваться, так как в тонком кишечнике переваривание белков происходит чрезвычайно эффективно.
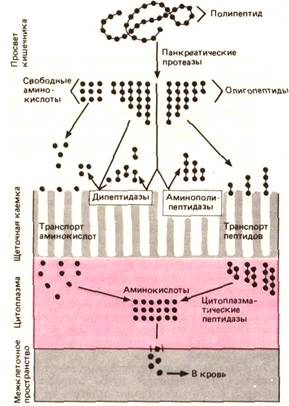
Через 10-20 мин после приема пищи начинается образованиепанкреатических пептидаз, которое продолжается до тех пор, пока белки присутствуют в кишечнике. Часть этих ферментов выделяется с калом. Содержание химотрипсина в каловых массах может служить показателем функциональной активности поджелудочной железы. Различные панкреатические ферменты, катализирующие гидролиз белков, атакуют белковую молекулу в разных участках (табл. 29.2). В двенадцатиперстной кишке, куда эти ферменты поступают в неактивной форме, происходит их активация трипсином, образующимся из трипсиногена под действием энтерокиназы. Ферменты, гидролизующие белки, подразделяют наэндопептидазы (трипсин, химотрипсин, эластаза) иэкзопептидазы (карбопептидазы А и В).  Эндопептидазы расщепляют внутренние связи в белковой молекуле с образованием олигопептидов. Экзопептидазы отщепляют аминокислоты с концов пептидной цепи. Около 30% конечных продуктов гидролиза составляют нейтральные и основные аминокислоты и 70% - олигопептиды, состоящие из 2-6 аминокислотных остатков. В щеточной каемке и внутри энтероцитов присутствуют другие пептидазы. В цитозоле подвергается гидролизу примерно 90% олигопептидов (ди- и трипептиды), поступающих в клетку при участии специальных транспортных систем. Около 10% олигопептидов, главным образом состоящих из 4-8 аминокислотных остатков, гидролизуют ферменты, локализованные в щеточной каемке. В каждом случае итог процесса - появление в крови воротной вены аминокислот как конечных продуктов гидролитического расщепления белков.
Всасывание белков, пептидов и аминокислот. Примерно 50-60% белков пищи всасывается в двенадцатиперстной кишке и около 30% - по мере прохождения химуса до подвздошной кишки, т.е. 80-90% экзогенных и эндогенных белков всасывается в тонком кишечнике. Только около 10% белков достигает толстого кишечника, где они расщепляются под действием бактерий. Небольшое количество белка выделяется с калом, но эта часть белка приходится на слущивающиеся клетки, а не на непереваренные остатки пищи. Интактные молекулы белка поглощаются в очень небольшом количестве путем пиноцитоза (с. 751). Всасывание по этому пути не имеет значения для усвоения белков, но может играть важную роль в связи с иммунореактивностью, приводя к сенсибилизации и аллергии. Пептиды всасываются в виде ди- и трипептидов путем пассивного переноса или активного транспорта с участием переносчиков.
Поглощение аминокислот происходит с помощью четырех основных групп транспортных систем: для 1. нейтральных, 2. двухосновных 3. дикарбоновых аминокислот 4. иминокислот. 5. К дополнительной группе относится система для глицина. Системы первых трех групп осуществляют перенос по механизму сопряжения с транспортом Na+ описанному выше (вторичноактивный транспорт) (с. 773); при этом всасывание кислых дикарбоновых аминокислот происходит как пассивный процесс, хотя и с участием переносчиков. За счет внутриклеточного переаминирования с участием аланина концентрация этих аминокислот в клетке сохраняется низкой. Различные аминокислоты одной группы ингибируют перенос друг друга, конкурируя за один и тот же переносчик (конкурентное ингибирование).
Рис. 29.37. Переваривание и всасывание белков. Просвет кишечника: расщепление полипептидов до олигопептидов, ди- и трипептидов и аминокислот. Мембраны клеток щеточной каемки: дальнейшее расщепление специфическими пептидазами и поглощение аминокислот и олигопептидов. Цитоплазма: расщепление ди- и олигопептидов цитоплазматическими пептидазами до аминокислот. Базальная мембрана: выход аминокислот из клетки в кровь
Нуклеопротеины гидролизуются и всасываются так же, как другие белки. Нуклеиновые кислоты - ДНК и РНК - гидролизуются специальными панкреатическими ферментами - дезоксирибонуклеаэой и рибонуклеазой и расщепляются в щеточной каемке фосфодиэстеразами и нуклеотидазами до нуклеотидов. Нуклеотиды транспортируются в энтероциты при участии специальных механизмов (рис. 29.37). Переваривание и всасывание ЖИРОВ
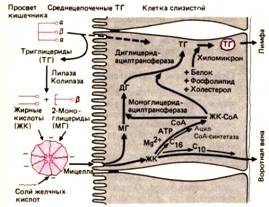
Человек потребляет в день около 60 -100 г жиров. Примерно 90% жиров пищи - это триглицериды, большую часть которых составляют липиды, содержащие жирные кислоты с длинной цепью из 16 (пальмитиновая кислота) или 18 (стеариновая, олеиновая, линолевая кислоты) атомов углерода. Триглицериды, содержащие жирные кислоты с короткой цепью (2-4 углеродных атома) или средней цепью (6-8 атомов), составляют лишь небольшую часть жиров пищи. Остальные 10% жиров пищи приходятся на фосфолипиды (главным образом лецитин), эфиры холестерола и жирорастворимые витамины (рис. 29.38). Переваривание жиров. В желудке жиры образуют капельки диаметром около 100 нм. В щелочной среде тонкого кишечника при наличии белков, продуктов расщепления предшествующей порции жиров, лецитина и желчных кислот жиры образуют эмульсию с размером капелек около 5 нм. В тонком кишечнике жиры стимулируют выделение клетками слизистой холецистокинина, активирующего секрецию ферментов поджелудочной железы и сокращения желчного пузыря, Липаза, секретируемая поджелудочной железой,состоит из двух компонентов – колипазы, образующейся в результате активации проколипазы трипсином и локализованной на границе раздела водной и липидной фаз, и панкреатической липазы, образующей комплекс с колипазой. [1] Липаза катализирует отщепление от триглицеридов жирных кислот в положениях 1 и 3 с. Конечный продукт — жирные кислоты, диацилглицерины и моноацилглицерины. Количество липазы, поступающей с панкреатическим соком, так велико, что к тому моменту, когда жир достигает середины двенадцатиперстной кишки, 80% его оказывается гидролизованным. В связи с этим нарушение переваривания жиров, связанное с недостаточностью липазы, не выявляется вплоть до полного прекращения деятельности поджелудочной железы или сильного ее разрушения. Помимо липазы поджелудочная железа секретирует и другие ферменты липидного обмена, также активируемые трипсином. К числу этих ферментов относитсяфосфолипаза Ад, которая в присутствии ионов Са2+ и желчных кислот отщепляет жирную кислоту от фосфолипида лецитина с образованием лизолецитина. Холестерол обычно присутствует в пище в виде эфиров и высвобождается под действием холестеролэстеразы.
Рис.29.38. Переваривание и всасывание липидов. В просвете кишечника триглицериды расщепляются под действием колипазы и липазы до жирных кислот и 2-моноглицеридов, которые содержатся в растворе в виде мицелл и поступают из них в энтероциты. В клетках из длинноцепочечных жирных кислот и 2-моноглицеридов ресинтезируются триглицериды, которые в виде заключенных в белковую оболочку хиломикронов выходят в лимфу. Жирные кислоты с короткими или средними цепями поглощаются и переносятся в кровь непосредственно в этой форме. МГ-моноглицериды, ДГ-диглицериды, ТГ-триглицериды, ЖК-жирные кислоты (по [27] с изменениями)
Продукты гидролиза липидов плохо растворимы в воде и могут находиться в кишечнике в растворенном виде лишь в составе мицелл (с. 767). Простые мицеллы, состоящие только из желчных кислот (чистые мицеллы), после внедрения в их гидрофобную сердцевину жирных кислот, моноглицеридов, фосфолипидов и холестерола превращаются всмешанные мицеллы. Благодаря растворимости этих мицелл в воде концентрация конечных продуктов гидролитического расщепления липидов в просвете кишечника повышается в тысячи раз. Жирные кислоты с короткими и средними цепями и содержащие их липиды довольно хорошо растворимы в воде и могут диффундировать к поверхности энтероцитов, не встраиваясь в мицеллы. Всасывание продуктов гидролитического расщепления жиров Жиры всасываются так эффективно, что 95% триглицеридов (но лишь 20-50% холестерола) поглощается уже из просвета двенадцатиперстной кишки и верхнего отдела тощей кишки. У человека при обычной диете в сутки выделяется с калом до 5-7 г жира. При безжировой диете эта величина уменьшается до 3 г/сут, а источником жира служат слущивающиеся эпителиальные клетки и бактерии. Прежде чем попасть внутрь энтероцита, компонентысмешанных мицелл должны преодолеть три барьера: 1) неперемешивающийся водный слой, прилежащий к поверхности клетки, - основное препятствие для жирных кислот с длинными цепями и моноглицеридов и для выполнения мицеллами их функций; 2) слой слизи, покрывающий щеточную каемку; при толщине 2-4 мкм этот слой также препятствует переносу компонентов мицелл; 3) липидную мембрану энтероцита.
Мицеллы в клетку не проникают, но их липидные компоненты растворяются в плазматической мембране и быстро диффундируют в клетку по концентрационному градиенту. Остаточное вещество мицелл может затем возвратиться в просвет и включить новые липидные компоненты. Внутриклеточный синтез липидов Пройдя через клеточную поверхность, продукты расщепления жиров - моноглицериды и жирные кислоты - транспортируются небольшим белком с мол. массой 12000 в микросомы эндоплазматического ретикулума, где из них вновь синтезируются триглицеридыи другие липиды. Вначале при участии моноглицеридтрансферазы образуются эфиры активированных жирных кислот с моноглицеридами, а затем при участии диглицеридтрансферазы к синтезированному диглицериду присоединяется еще одна жирная кислота и образуется триглицерид. Другой, менее важный путь ресинтеза триглицеридов связан с обменом глюкозы. Фосфолипиды, как и триглицериды, могут образовываться в энтероцитах путем этерификации (примером служит образование лецитина из лизолецитина). Реэтерификация холестерола катализируется холестеролэстеразой. Кроме того, в подвздошной кишке происходит синтез холестерола de novo, поэтому тонкому кишечнику принадлежит особая роль в обмене этого липида.
Образование хиломикронов Новообразованные триглицериды и другие липиды, прежде чем покинуть клетку, покрываются специальной оболочкой, содержащей холестерол и фосфолипиды в сочетании со специфическими гликопротеинами, синтезированными в аппарате Гольджи. Состав этих сложных образований, называемых хиломикронами, приблизительно следующий: 90%-триглицериды, 7%-фосфолипиды, 2%-холестерол и 1%-белок. Диаметр хиломикронов составляет 60 – 75 нм в зависимости от скорости всасывания и ресинтеза жиров. Хиломикроны накапливаются в секреторных везикулах; последние сливаются с боковой мембраной клетки, и через образующееся при этом отверстие хиломикроны выходят в межклеточное пространство, откуда по центральному лимфатическому и грудному протокам поступают в кровь. После приема жирной пищи содержание хиломикронов в плазме повышается так сильно, что она становится по цвету молочно-белой (алиментарная гиперлипемия). Известно врожденное заболевание - А-β-липопротеинемия, - связанное с нарушением высвобождения хиломикронов из клеток. При этом заболевании поглощенный жир не выводится из клетки и остается в ней. Помимо хиломикронов в энтероцитах образуются липопротеины очень низкой плотности, которые также высвобождаются в лимфатические сосуды. |
||
|
|
Последнее изменение этой страницы: 2018-05-10; просмотров: 334. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |