|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
Моноклональные антитела. Получение. Использование.Моноклональные антитела (МКА) – это иммуноглобулины, вырабатываемые иммунными клетками, принадлежащими к одному клеточному клону.Гибридо́ма — гибридная клеточная линия, полученная в результате слияния клеток двух видов: способных к образованию антител B-лимфоцитов, полученных из селезёнки иммунизированного животного (чаще всего мыши), и опухолевых клеток миеломы. Слияние клеток производится с помощью нарушающего мембраны агента, такого, как полиэтиленгликоль или вирус Сёндай. Поскольку раковые клетки миеломы «бессмертны», то есть способны делиться большое количество раз, после слияния и соответствующей селекции гибридома, производящая моноклональные антитела против антигена может поддерживаться долгое время. В 1984 г. за открытие принципа получения моноклональных антител Мильштейн, Кёлер и Ерне получили Нобелевскую премию по физиологии и медицине.
(1) иммунизация животных; (2) выделение В-лимфоцитов из селезенки; (3) культура клеток миеломы; (4) слияние В-лимфоцитов и клеток миеломы; (5) сегрегация (отделение) клеточных линий; (6) скрининг и селекция линий, производящих антитела; (7) размножениегибридомы in vitro (a) или in vivo (b); (8) получение антител (invivo путём введения клеточной суспензии в брюшную полость мышей, где гибридома размножается подобно раковым клеткам материнской миеломы, секретируя антитела во внутриполостную жидкость с образованием асцита либо в биореакторе invitro)
Нагрузочные серологические реакции. Реакции непрямой гемагглютинации. Компоненты. Применение. Это реакции между антигеном и антителом, которые происходят в живом организме, а также могут быть воспроизведены в лабораторных условиях. Реакции антитела и антигена называются серологическими, или гуморальными. В процессе взаимодействия с антигеном участвует не вся молекула иммуноглобулина, а лишь ее ограниченный участок — антигенсвязывающий центр. Антитело взаимодействует не со всей молекулой антигена сразу, а только с ее антигенной детерминантой. Антитела обладают специфичностью взаимодействия, т. е. связываются со строго определенной антигенной детерминантой.  При выделении микроорганизмов от больного в бактериологических лабораториях проводят идентификацию возбудителей путем изучения их антигенных свойств с помощью иммунных диагностических сывороток, содержащих специфические антитела. В микробиологической практике широко применяются реакции агглютинации, преципитации, нейтрализации и т. д. Эти реакции различаются по технике постановки, хотя все они основаны на реакции взаимодействия антигена с антителом и применяются для выявления как антител, так и антигенов. Реакция непрямой (пассивной) гемагглютинации (РНГА, РПГА) основана на использовании эритроцитов (или латекса) с адсорбированными на их поверхности антигенами или антителами, взаимодействие которых с соответствующими антителами или антигенами сыворотки крови больных вызывает склеивание и выпадение эритроцитов на дно пробирки или ячейки в виде фестончатого осадка. При обнаружении антигенов реакция обозначается обратной пассивной гемагглютинацией - РОПГА (reversed passive hemagglutination test), а при обнаружении антител - пассивной гемагглютинацией - РПГА (passive hemagglutination test). РНГА нашла широкое применение для выявления серологических маркеров инфицирования вирусом гепатита В, особенно HBsAg и антител к нему. Компоненты. Для постановки РНГА могут быть использованы эритроциты барана, лошади, кролика, курицы, мыши, человека и другие, которые заготавливают впрок, обрабатывая формалином или глютаральдегидом. Адсорбционная емкость эритроцитов увеличивается при обработке их растворами танина или хлорида хрома. Антигенами в РНГА могут служить полисахаридные АГ микроорганизмов, экстракты бактериальных вакцин, АГ вирусов и риккетсий, а также другие вещества. Эритроциты, сенсибилизированные АГ, называются эритроцитарными диагностикумами. Для приготовления эритроцитарного диагностикума чаще всего используют эритроциты барана, обладающие высокой адсорбирующей активностью. Применение. РНГА применяют для диагностики инфекционных болезней, определения гонадотропного гормона в моче при установлении беременности, для выявления повышенной чувствительности к лекарственным препаратам, гормонам и в некоторых других случаях. Механизм. Реакция непрямой гемагглютинации (РНГА) отличается значительно более высокой чувствительностью и специфичностью, чем реакция агглютинации. Ее используют для идентификации возбудителя по его антигенной структуре или для индикации и идентификации бактериальных продуктов — токсинов в исследуемом патологическом материале. Соответственно используют стандартные (коммерческие) эритроцитарные антительные диагностикумы, полученные путем адсорбции специфических антител на поверхности танизированных (обработанных танином) эритроцитов. В лунках пластмассовых пластин готовят последовательные разведения исследуемого материала. Затем в каждую лунку вносят одинаковый объем 3 % суспензии нагруженных антителами эритроцитов. При необходимости реакцию ставят параллельно в нескольких рядах лунок с эритроцитами, нагруженными антителами разной групповой специфичности. Через 2 ч инкубации при 37 °С учитывают результаты, оценивая внешний вид осадка эритроцитов (без встряхивания): при отрицательной реакции появляется осадок в виде компактного.диска или кольца на дне лунки, при положительной реакции — характерный кружевной осадок эритроцитов, тонкая пленка с неровными краями.
Наследственность и изменчивость у бактерий. Механизмы обеспечения и передачи дочерним клеткам генетической информации. Механизмы изменчивости.
У микроорганизмов, как и у других живых существ, наблюдается наследование признаков, свойственных определенному виду. В опытах было показано, что, если содержать бактерии в определенных постоянных условиях, они длительно сохраняют свои свойства. Это говорит о стабильности признаков, присущих каждому виду бактерий. Потомство микробной клетки в основном наследует ее свойства, что позволяет определить, идентифицировать, любой вид микроорганизмов. Однако известно, что у одного и того же вида бактерий возможны отклонения морфологических и физиологических свойств, возникающие под влиянием факторов внешней среды. Например, Пастер искусственным путем получил необратимые изменения у возбудителей сибирской язвы и бешенства и создал вакцины, предохраняющие от этих заболеваний. Н. Ф. Гамалея наблюдал изменения морфологии холерного вибриона при добавлении в питательную среду хлорида лития. Это свидетельствует о том, что микроорганизмы могут изменять свои свойства в зависимости от условий существования. У микроорганизмов различают фенотипическую изменчивость — ненаследуемую и генотипическую изменчивость — наследуемую, которые связаны с двумя основными особенностями клетки: ее генотипом и фенотипом. Генотип — общая сумма генов, которыми обладает клетка. Он определяет целую группу свойств организма, специфичных для данного вида микроорганизмов, которые могут по разному проявиться в различных условиях внешней среды. Конъюгация бактерий состоит в переходе генетического материала (ДНК) из клетки-донора («мужской») в клетку-реципиент («женскую») при контакте клеток между собой. Мужская клетка содержит F-фактор, или половой фактор, который контролирует синтез так называемых половых пилей, или F-пилей. Клетки, не содержащие F-фактора, являются женскими; при получении F-фактора они превращаются в «мужские» и сами становятся донорами. F-фактор располагается в цитоплазме в виде кольцевой двунитчатой молекулы ДНК, т. е. является плазмидой. Молекула F-фактора значительно меньше хромосомы и содержит гены, контролирующие процесс конъюгации, в том числе синтез F-пилей. При конъюгации F-пили соединяют «мужскую» и «женскую» клетки, обеспечивая переход ДНК через конъюгационный мостик или F-пили. Клетки, содержащие F-фактор в цитоплазме, обозначаются F+; они передают F-фактор клеткам, обозначаемым F" («женским»), не утрачивая донорской способности, так как оставляют копии F-фактора. Если F-фактор включается в хромосому, то бактерии приобретают способность передавать фрагменты хромосомной ДНК и называются Hfr-клетками (от англ. highfrequencyofrecombination — высокая частота рекомбинаций), т.е. бактериями с высокой частотой рекомбинаций. При конъюгации клеток Hfr и клеток F" хромосома разрывается и передается с определенного участка (начальной точки) в клетку F", продолжая реплицироваться. Перенос всей хромосомы может длиться до 100 мин. Переносимая ДНК взаимодействует с ДНК реципиента — происходит гомологичная рекомбинация. Прерывая процесс конъюгации бактерий, можно определять последовательность расположения генов в хромосоме. Иногда F-фактор может при выходе из хромосомы захватывать небольшую ее часть, образуя так называемый замещенный фактор — F'. При конъюгации происходит только частичный перенос генетического материала, поэтому ее не следует отождествлять полностью с половым процессом у других организмов. Трансдукция — передача ДНК от бактерии-донора к бактерии-реципиенту при участии бактериофага. Различают неспецифическую (общую) трансдукцию, при которой возможен перенос любого фрагмента ДНК донора, и специфическую — перенос определенного фрагмента ДНК донора только в определенные участки ДНК реципиента. Неспецифическая трансдукция обусловлена включением ДНК донора в головку фага дополнительно к геному фага или вместо генома фага (дефектные фаги). Специфическая трансдукция обусловлена замещением некоторых генов фага генами хромосомы клетки-донора. Фаговая ДНК, несущая фрагменты хромосомы клетки-донора, включается в строго определенные участки хромосомы клетки-реципиента. Таким образом, привносятся новые гены и ДНК фага в виде профага репродуцируется вместе с хромосомой, т.е. этот процесс сопровождается лизоге-нией. Если фрагмент ДНК, переносимый фагом, не вступает в рекомбинацию с хромосомой реципиента и не реплицируется, но с него считывается информация о синтезе соответствующего продукта, такая трансдукция называется абортивной. Трансформация заключается в том, что ДНК, выделенная из бактерий в свободной растворимой форме, передается бактерии-реципиенту. При трансформации рекомбинация происходит, если ДНК бактерий родственны друг другу. В этом случае возможен обмен гомологичных участков собственной и проникшей извне ДНК. Впервые явление трансформации описал Ф. Гриффите (1928). Он вводил мышам живой невирулентный бескапсульный R-штамм пневмококка и одновременно убитый вирулентный капсульный S-штамм пневмококка. Из крови погибших мышей был выделен вирулентный пневмококк, имеющий капсулу убитого S-штамма пневмококка. Таким образом, убитый S-штамм пневмококка передал наследственную способность капсулообразования R-штамму пневмококка. О. Эвери, К. Мак-Леод и М. Мак-Карти (1944) доказали, что трансформирующим агентом в этом случае является ДНК. Путем трансформации могут быть перенесены различные признаки: капсулообразование, устойчивость к антибиотикам, синтез ферментов. Изучение бактериальной трансформации позволило установить роль ДНК как материального субстрата наследственности. При изучении генетической трансформации у бактерий были разработаны методы экстракции и очистки ДНК, биохимические и биофизические методы ее анализа. Плазмиды — внехромосомные мобильные генетические структуры бактерий, представляющие собой замкнутые кольца двунитчатой ДНК. По размерам составляют 0,1—5 % ДНК хромосомы. Плазмиды способны автономно копироваться (реплицироваться) и существовать в цитоплазме клетки, поэтому в клетке может быть несколько копий плазмид. Плазмиды могут включаться (интегрировать) в хромосому и реплицироваться вместе с ней. Различают трансмиссивные и нетрансмиссивные плазмиды. Трансмиссивные (конъюгативные) плазмиды могут передаваться из одной бактерии в другую. Среди фенотипических признаков, сообщаемых бактериальной клетке плазмидами, можно выделить следующие: 1) устойчивость к антибиотикам; 2) образование колицинов; 3) продукция факторов патогенности; 4) способность к синтезу антибиотических веществ; 5) расщепление сложных органических веществ; 6) образование ферментов рестрикции и модификации. Плазмиды могут определять вирулентность бактерий, например возбудителей чумы, столбняка, способность почвенных бактерий использовать необычные источники углерода, контролировать синтез белковых антибиотикоподобных веществ — бактериоцинов, детерминируемых плазмидами бактериоциногении, и т. д. Существование множества других плазмид у микроорганизмов позволяет полагать, что аналогичные структуры широко распространены у самых разнообразных микроорганизмов. Плазмиды подвержены рекомбинациям, мутациям, могут быть элиминированы (удалены) из бактерий, что, однако, не влияет на их основные свойства. Плазмиды являются удобной моделью для экспериментов по искусственной реконструкции генетического материала, широко используются в генетической инженерии для получения рекомбинантных штаммов. Благодаря быстрому самокопированию и возможности конъюгаци-онной передачи плазмид внутри вида, между видами или даже родами плазмиды играют важную роль в эволюции бактерий. Неспецифические факторы защиты организма. Фагоцитоз. Показатели фагоцитоза. |
||
|
|
Последнее изменение этой страницы: 2018-04-12; просмотров: 747. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |
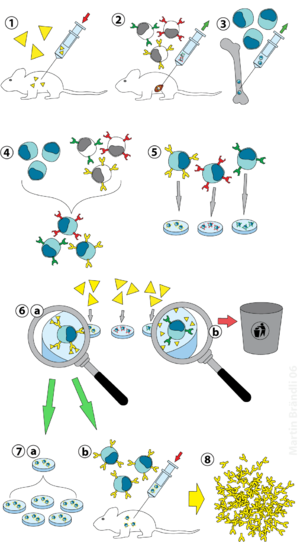 Гибридомная технология:
Гибридомная технология: