|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
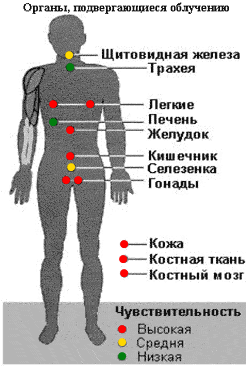
Вороний глаз - ParisguadrifoliaL., Ландышмайский - Convallaria majalis L.,Лютикедкий - Ranunculus acer L. 4,С- личинка трихинелла спиралис. Билет 95 В1. 81.Организация генома эукариот. У эукариотических организмов механизм регуляции транскрипции гораздо более сложен. В результате клонирования и секвенирования генов эукариот обнаружены специфические последовательности, принимающие участие в транскрипции и трансляции. Для эукариотической клетки характерно: 1. Наличие интронов и экзонов в молекуле ДНК. 2. Созревание и-РНК - вырезание интронов и сшивка экзонов. 3. Наличие регуляторных элементов, регулирующих транскрипцию, таких как: а) промоторы - 3 вида, на каждый из которых садится специфическая полимераза. Pol I реплицирует рибосомные гены, Pol II - структурные гены белков, Pol III - гены, кодирующие небольшие РНК. Промотор Pol I и Pol II находятся перед участком инициации транскрипции, промотор Pol III - в рамках структурного гена; б) модуляторы - последовательности ДНК, усиливающие уровень транскрипции; в) усилители - последовательности, усиливающие уровень транскрипции и действующие независимо от своего положения относительно кодирующей части гена и состояния начальной точки синтеза РНК; г) терминаторы - специфические последовательности, прекращающие и трансляцию, и транскрипцию. Эти последовательности по своей первичной структуре и расположению относительно инициирующего кодона отличаются от прокариотических, и бактериальная РНК-полимераза их не "узнает". Таким образом, для экспрессии эукариотических генов в клетках прокариот нужно, чтобы гены находились под контролем прокариотических регуляторных элементов. Это обстоятельство необходимо учитывать при конструировании векторов для экспрессии.  В2. ---- В3. 21.О влиянии радиации на организм человека. То, что радиация оказывает пагубное влияние на здоровье человека, уже ни для кого не секрет. Когда радиоактивное излучение проходит через тело человека или же когда в организм попадают зараженные вещества, то энергия волн и частиц передается нашим тканям, а от них клеткам. В результате атомы и молекулы, составляющие организм, приходят в возбуждение, что ведёт к нарушению их деятельности и даже гибели. Все зависит от полученной дозы радиации, состояния здоровья человека и длительности воздействия. Для ионизирующего излучения нет барьеров в организме, поэтому любая молекула может подвергнуться радиоактивному воздействию, последствия которого могут быть самыми разнообразными. Возбуждение отдельных атомов может привести к перерождению одних веществ в другие, вызвать биохимические сдвиги, генетические нарушения и т.п. Пораженными могут оказаться белки или жиры, жизненно необходимые для нормальной клеточной деятельности. Таким образом, радиация воздействует на организм на микроуровне, вызывая повреждения, которые заметны не сразу, а проявляют себя через долгие годы. Поражение отдельных групп белков, находящихся в клетке, может вызвать рак, а также генетические мутации, передающиеся через несколько поколений. Воздействие малых доз облучения обнаружить очень сложно, ведь эффект от этого проявляется через десятки лет.
Билет 96 В1. 88.Цитологические основы полового размножения. Мейоз, как специфический процесс при формировании половых клеток. Мейоз (от греч.meiosis — уменьшение) или редукционное деление клетки — деление ядра эукариотическойклетки с уменьшением числа хромосом в два раза. Происходит в два этапа (редукционный и эквационный этапы мейоза). Мейоз не следует смешивать с гаметогенезом — образованием специализированных половых клеток, или гамет, из недифференцированныхстволовых. С уменьшением числа хромосом в результате мейоза в жизненном цикле происходит переход от диплоидной фазы к гаплоидной. Восстановление плоидности (переход от гаплоидной фазы к диплоидной) происходит в результате полового процесса. В связи с тем, что в профазе первого, редукционного, этапа происходит попарное слияние (конъюгация) гомологичных хромосом, правильное протекание мейоза возможно только в диплоидных клетках или в чётных полиплоидах (тетра-, гексаплоидных и т. п. клетках). Мейоз может происходить и в нечётных полиплоидах (три-, пентаплоидных и т. п. клетках), но в них, из-за невозможности обеспечить попарное слияние хромосом в профазе I, расхождение хромосом происходит с нарушениями, которые ставят под угрозу жизнеспособность клетки или развивающегося из неё многоклеточного гаплоидного организма. Этот же механизм лежит в основе стерильности межвидовых гибридов. Поскольку у межвидовых гибридов в ядре клеток сочетаются хромосомы родителей, относящихся к различным видам, хромосомы обычно не могут вступить в конъюгацию. Это приводит к нарушениям в расхождении хромосом при мейозе и, в конечном счете, к нежизнеспособности половых клеток, или гамет. Определенные ограничения на конъюгацию хромосом накладывают и хромосомные мутации (масштабные делеции, дупликации, инверсии или транслокации). Мейоз состоит из 2 последовательных делений с короткой интерфазой между ними.
· Лептотена или лептонема — упаковка хромосом. · Зиготена или зигонема — конъюгация (соединение) гомологичных хромосом с образованием структур, состоящих из двух соединённых хромосом, называемых тетрадами или бивалентами. · Пахитена или пахинема — кроссинговер (перекрест), обмен участками между гомологичными хромосомами; гомологичные хромосомы остаются соединенными между собой. · Диплотена или диплонема — происходит частичная деконденсация хромосом, при этом часть генома может работать, происходят процессы транскрипции (образование РНК), трансляции (синтез белка); гомологичные хромосомы остаются соединёнными между собой. У некоторых животных в ооцитах хромосомы на этой стадии профазы мейоза приобретают характерную форму хромосом типа ламповых щёток. · Диакинез — ДНК снова максимально конденсируется, синтетические процессы прекращаются, растворяется ядерная оболочка; центриоли расходятся к полюсам; гомологичные хромосомы остаются соединёнными между собой.
Второе деление мейоза следует непосредственно за первым, без выраженной интерфазы: S-период отсутствует, поскольку перед вторым делением не происходит репликации ДНК.
В результате из одной диплоидной клетки образуется четыре гаплоидных клетки. В тех случаях, когда мейоз сопряжён с гаметогенезом (например, у многоклеточных животных), при развитии яйцеклеток первое и второе деления мейоза резко неравномерны. В результате формируется одна гаплоидная яйцеклетка и два так называемых редукционных тельца (абортивные дериваты первого и второго делений). В2. 19.Классификация генов человека по структуре и функциям. Ген представляет собой последовательность нуклеотидов ДНК размером от нескольких сотен до миллиона пар нуклеотидов, в которых закодирована генетическая информация о первичной структуре белка (число и последовательность аминокислот). Для регулярного правильного считывания информации в гене должны присутствовать: кодон инициации, множество смысловых кодонов и кодон терминации. Три подряд расположенных нуклеотида представляют собой кодон, который и определяет, какая аминокислота будет располагаться в данной позиции в белке. Например, в молекуле ДНК последовательность оснований ТАС является кодоном для аминокислоты метионина, а последовательность ТТТ кодирует фенилаланин. В молекуле иРНК вместо тимина (Т) присутствует основание урацил (У). Таблица генетического кода во всех руководствах представлена именно символами иРНК. Из 64 возможных кодонов смысловыми являются 61, а три триплета — УАА, УАГ, УГА — не кодируют аминокислоты и поэтому были названы бессмысленными, однако на самом деле они представляют собой знаки терминации трансляции. В3. 23.Факторы, влияющие на изменение климата. Изменение климата — колебания климатаЗемли в целом или отдельных её регионов с течением времени, выражающиеся в статистически достоверных отклонениях параметров погоды от многолетних значений за период времени от десятилетий до миллионов лет. Учитываются изменения как средних значений погодных параметров, так и изменения частоты экстремальных погодных явлений. Изучением изменений климата занимается наука палеоклиматология. Причиной изменения климата являются динамические процессы на Земле, внешние воздействия, такие как колебания интенсивности солнечного излучения, и, по одной из версий, с недавних пор, деятельность человека. В последнее время термин «изменение климата» используется как правило (особенно в контексте экологической политики) для обозначения изменения в современном климате (см. глобальное потепление). Изменения климата обусловлены переменами в земной атмосфере, процессами, происходящими в других частях Земли, таких как океаны, ледники, а также эффектами, сопутствующими деятельности человека. Внешние процессы, формирующие климат, — это изменения солнечной радиации и орбиты Земли. |
||
|
|
Последнее изменение этой страницы: 2018-04-12; просмотров: 501. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |
 α D
α D 
 A B
A B  C
C