|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
Определение Х- и Y-хроматинаСтр 1 из 25Следующая ⇒ Развитие половых клеток. Развитие половых клеток называется прогенезом, или гаметогенезом, и происходит в половых железах. Суть гаметогенеза состоит в образовании из диплоидных стволовых предшественников половых клеток высокодифференцированных клеток (сперматозоидов и яйцеклеток) с гаплоидным набором хромосом. Развитие мужских половых клеток — сперматогенез, включает четыре фазы: размножения, роста, созревания, формирования. В результате сперматогенеза возникают клетки сперматозоиды (спермин), содержащие Х- или Y-половую хромосому. Развитие женских половых клеток — овогенез, состоит из трех фаз: размножения, роста, созревания. В результате овогенеза образуются яйцеклетки (овоциты), содержащие Х-половую хромосому. В основном мужской и женский гаметогенезы протекают однотипно. В фазе размножения исходные половые клетки (сперматогонии — в сперматогенезе и овогонии — в овогенезе) в ходе клеточного цикла синтезируют ДНК и митотически делятся; их размножение протекает на основе общих закономерностей, характерных и для соматических клеток. В процессе сперматогенеза делится только часть сперматогонии, так называемые светлые сперматогонии типа А, которые являются полустволовыми клетками, дающими начало сперматогониям типа В. Сперматогонии типа В проходят несколько митотических циклов, при этом не происходит полной цитотомии сперматогонии и они остаются связанными цитоплазматическими мостиками. Фаза размножения сперматогонии контролируется естественными митогенами — гормоном аденогипофиза фоллитропином и интерлейкином-1, вырабатываемым клетками Сертоли и клетками Лейдига (интерстициальными эндокриноцитами) яичек.  В2.Методы в генетике человека. Биохимический метод. Дерматоглифика. В отличие от цитогенетического метода, который позволяет изучать структуру хромосом и кариотипа в норме и диагностировать наследственные болезни, связанные с изменением их числа и нарушением организации, наследственные заболевания, обусловленные генными мутациями, а также полиморфизм по нормальным первичным продуктам генов изучают с помощью биохимических методов. Впервые эти методы стали применять для диагностики генных болезней еще в начале XX в. В последние 30 лет их широко используют в поиске новых форм мутантных аллелей. С их помощью описано более 1000 врожденных болезней обмена веществ. Для многих из них выявлен дефект первичного генного продукта. Наиболее распространенными среди таких заболеваний являются болезни, связанные с дефектностью ферментов, структурных, транспортных или иных белков. Дефекты структурных и циркулирующих белков выявляются при изучении их строения. Так, в 60-х гг. XX в. был завершен анализ (3-глобино-вой цепи гемоглобина, состоящей из 146 аминокислотных остатков. Установлено большое разнообразие гемоглобинов у человека, связанное с изменением структуры его пептидных цепей, что нередко является причиной развития заболеваний (см. § 4.1). Дефекты ферментов устанавливают путем определения содержания в крови и моче продуктов метаболизма, являющихся результатом функционирования данного белка. Дефицит конечного продукта, сопровождающийся накоплением промежуточных и побочных продуктов нарушенного метаболизма, свидетельствует о дефекте фермента или его дефиците в организме (см. § 4.1). Биохимическую диагностику наследственных нарушений обмена проводят в два этапа. На первом этапе отбирают предположительные случаи заболеваний, на втором —более точными и сложными методами уточняют диагноз заболевания. Применение биохимических исследований для диагностики заболеваний в пренатальном периоде или непосредственно после рождения позволяет своевременно выявить патологию и начать специфические медицинские мероприятия, как, например, в случае фенилкетонурии. Для определения содержания в крови, моче или амниотической жидкости промежуточных, побочных и конечных продуктов обмена кроме качественных реакций со специфическими реактивами на определенные вещества используют хроматографические методы исследования аминокислот и других соединений. В3.Популяционный уровень взаимодействия паразитов и хозяев. Типы регуляции и механизмы устойчивости системы «паразит-хозяин». С представлением о паразите неразрывно связано понятие о его хозяине. Хозяин паразита - это организм животного или человека, в теле которого обитает паразит временно или постоянно и размножается половым или бесполым путем. Смена хозяина обусловлена стадийностью жизненного цикла паразита. Личиночные стадии, как правило, развиваются в организме одного вида, а половозрелые - другого вида. Перемена хозяев паразитами, в основном, обусловлена чередованием поколений, размножающихся половым или бесполым способами. Хозяин, в теле которого паразит достигает половозрелой стадии и размножается половым путем, называется 2 0окончательным или дефинитивным 2 0хозяином. Так, человек является окончательным хозяином для большинства видов цестод, трематод, нематод. Хозяин, в теле которого обитает личиночная стадия паразита, получил название 2 0промежуточного. Человек является промежуточным хозяином для малярийных плазмодиев, эхинококка, альвеококка и др. Для некоторых паразитов (описторхис, парагонимус, широкий лентец и др.) для завершения цикла развития требуется не один, а два (или несколько) промежуточных хозяев. Второго промежуточного хозяина называют дополнительным. Так, у описторхиса два промежуточных хозяина: первый - моллюск Bithynia leachi, дополнительный многие виды карповых рыб (плотва, линь, язь, карп, лещ, вобла, сазан и др.). У многих паразитов перемена хозяев не связана с чередованием поколений, но осуществляется по ходу развития одной и той же особи паразита. Например, степной клещ - переносчик клещевого сыпного тифа в Сибири (в состоянии личинки и нимфы нападает на различных мелких грызунов, а во взрослом состоянии - на крупных млекопитающих или человека). Практически один и тот же хозяин может иметь большое количество паразитов одного вида (малярийные плазмодии) или разных видов. У человека могут одновременно паразитировать острицы, власоглав, кишечные амебы, плазмодии и т.д. Хозяева, у которых паразиту обеспечивается наилучшая выживаемость, быстрый рост и наибольшая плодовитость, получили название облигатных, или 2 0обязательных. Человек является облигатным хозяином для дизентерийной амебы, аскариды, анкилостомы и других паразитов. Хозяин, в теле которого паразит может обитать, но не полностью адаптироваться, называется факультативным. Например, человек является облигатным хозяином для лентеца широкого. Однако эта цистода может паразитировать и в организме лисицы, но ее размеры при этом не достигают большой длины и срок жизни не превышает двух месяцев. Организм, в котором не происходит развитие паразита, а наблюдается лишь его накопление в инвазионной стадии, называется 2 0резервуарным хозяином. Последний кумулирует возбудителя и сохраняет инвазию, что способствует большей зараженности окончательного хозяина. Например, щука, поедая дополнительного хозяина широкого лентеца (карповых рыб), накапливает в своих тканях личиночные стадии (плероцеркоиды) паразита и таким образом выполняет функцию резервуарного хозяина. Билет 4 В1.Закономерности сперматогенеза у млекопитающих и человека. Семенник состоит из нескольких тысяч цилиндрических семенных канальцев, в каждом из которых развиваются миллионы сперматозоидов. Стенки этих канальцев выстланы первичными, неспециализированными половыми клетками — сперматогониями. В течение эмбрионального развития и в детском возрасте сперматогонии делятся митотически, давая начало дополнительным сперматогониям, что ведет к росту семенника. После наступления половой зрелости некоторые из сперматогониев вступают в сперматогенез, состоящий из серии превращений, ведущих к образованию зрелых сперматозоидов. Другие сперматогонии продолжают делиться митотически и производят новые сперматогонии для последующих сперматогенезов. У большинства диких животных имеется определенный период, приходящийся на весну или на осень, когда семенники увеличиваются в размерах и в них происходит сперматогенез. В промежутке же между этими периодами семенник имеет небольшие размеры и содержит только сперматогонии. У человека и у большинства домашних животных сперматогенез после достижения половой зрелости протекает круглый год. В2.Методы в генетике человека. Близнецовый метод. Исследование генетических закономерностей на близнецах. Монозиготными близнецами называют индивидов, выросших из одной зиготы, т. е. обладающих идентичными генотипами. Дизиготные близнецы развиваются вследствие оплодотворения двух яйцеклеток, имеющих с генетической точки зрения, 50 % общих генов, т. е. сходные как братья и сестры. При близнецовом методе исследования проводят сопоставление монозиготных близнецов с дизиготными, а также монозиготных близнецов. При диагностике близнецовости используют метод сходства (сопоставление по внешним морфологическим признакам - пигментация кожи, волос, форма носа, губ, ушных раковин и т. д.), а также анализируют некоторые моногенные признаки - эритро- и лейкоцитарные антигены, группа белков крови, группы крови и т. д. Какой-либо качественный признак (заболевание) может встречаться у обоих близнецов (конкордантность) или у одного из них (дискордантность). Сопоставление степени парной конкордантности у моно- и дизиготных близнецов дает приблизительный ответ на вопрос о соотношении роли наследственности и среды в развитии заболевания. Если доминирующую роль в возникновении заболевания играет наследственность, то конкордантность у монозиготных близнецов выше, чем у дизиготных. Если в развитии болезни определенную роль играют негененетические факторы, то один из монозиготных близнецов может быть больным, другой здоровым. Следует все же учитывать, что определение роли среды и наследственности с помощью близнецового метода в формировании того или иного заболевания является сложным вопросом и требует более детального рассмотрения. Поэтому здесь следует упомянуть о недостатках близнецового метода, имеющих много биологических и социальных причин. Классический близнецовый метод: в этом случае используется такая схема эксперимента, при которой выраженность исследуемого признака сопоставляется в парах и близнецов и оценивается уровень внутрипарного сходства партнеров. Метод контрольного близнеца: этот метод используется на выборках МЗ близнецов. Так как МЗ близнецы весьма сходны по многим признакам, то из партнеров МЗ пар можно составить две выборки, уравненные по большому числу параметров. Такие выборки используют для исследования влияния конкретных средовых воздействий на изменчивость признака. При этом отобранная часть близнецов (по одному из каждой пары) подвергается специфическому воздействию, другая же часть является контрольной группой. Поскольку в эксперименте участвуют генетически идентичные люди, то этот способ можно считать моделью для изучения воздействия различных средовых факторов на одного и того же человека. Лонгитюдное близнецовое исследование: в этом случае проводится длительное наблюдение одних и тех же близнецовых пар. Фактически это сочетание классического близнецового метода с лонгитюдным. Широко используется для изучения влияния средовых и генетических факторов в развитии. Метод близнецовых семей: является сочетанием семейного и близнецового метода. При этом исследуются члены семей взрослых близнецовых пар. Дети МЗ близнецов по генетической конституции являются как бы детьми одного человека. Метод широко используется при изучении наследственных причин ряда заболеваний. Исследование близнецов как пары: предполагает исследование специфических близнецовых эффектов и особенностей внутрипарных отношений. Используется как вспомогательный метод для проверки справедливости гипотезы о равенстве средовых условий для партнеров МЗ и ДЗ пар. Сопоставление близнецов с неблизнецвми. Также вспомогательный метод, позволяющий оценить существенность разницы между близнецами и неблизнецами. Если разница между близнецами и остальными людьми не является значимой, то близнецы и остальные люди относятся к одной генеральной выборке и, следовательно, результаты бллизнецовых исследований можно распространять на всю популяцию. Так, было отмечено некоторое отставание членов близнецовых пар в развитии от одиночнорожденных. Особенно эта разница заметна в раннем возрасте. Но сопоставление результатов исследования членов близнецовых пар, чей партнер умер в раннем детстве и одиночнорожденных не выявил существенной разницы в уровне развития. То есть особенности развития близнецов обусловлены не столько трудностями эмбрионального развития, сколько с особенностями воспитания близнецов как пары (семейные трудности при рождении близнецов, замкнутость близнецов в паре и т.п.). Таким образом, близнецы несколько отличаются от всей популяции, но с возрастом эта разница заметно сглаживается и близнецы по большей части становятся сопоставимы с остальной популяцией. Метод разлученных близнецов: Из-за особенностей развития ДЗ и МЗ пар близнецов классический близнецовый метод и его разновидности принято считать “нежесткими” экспериментами: в них невозможно однозначно разделить влияние генетических и средовых факторов, так как в силу ряда причин условия развития близнецов по целому ряду причин оказываются несопоставимыми. Метод частично разлученных близнецов: Этот метод состоит в сравнении внутрипарного сходства МЗ и ДЗ близнецов, живущих какое-то время врозь. В этих исследованиях также можно определить в какой степени справедлив постулат о равенстве сред МЗ и ДЗ близнецов. Так, если МЗ близнецы живущие отдельно становятся все менее схожи друг с другом по некой психологической характеристике, а ДЗ близнецы, живущие врозь, не отличаются по внутрипарному сходству от вместе живущих ДЗ близнецов, то можно сделать вывод, что средовые условия МЗ и ДЗ неравноценны, а выводы о наследуемости изучаемой характеристики завышают показатель наследуемости этой характеристики. В3.Пути происхождения групп паразитов. Условия жизни паразита внутри тела своего хозяина резко отличаются от свободного существования во внешней среде. Эндопаразиту не приходится отыскивать себе пищу, у него нет необходимости в таких органах чувств, как глаза. Приспособление паразита к таким специальным условиям жизни неизбежно отражается на его организации и сводится к двум процесса: 1) к исчезновению тех органов, которые не требуются в новых условиях существования; 2) к возникновению, развитию или преобразованию органов, работа которых связана с особенностями жизни или питания на поверхности тела или внутри организма. Вся совокупность изменений организации паразитов сводится к упрощению строения и нередко к изменению внешнего вида. Паразитизм как явление живой природы возник у свободноживущих организмов в результате различных форм симбиоза и хищничества. Различают несколько категорий симбиоза: 1) мутуализм - такая форма сожительства, при которой оба партнера приносят друг другу какую либо пользу (лишайники - сожительство водорослей и грибов); 2) синойкия - сожительство, при котором один партнер использует другого в качестве временного убежища (рыба-горнак и двустворчатый моллюск беззубка); 3) коменсализм - такая категория сожительства, при которой один вид животного использует другого для питания остатками пищи другого вида (рыба-прилипала и акула); 4) паразитизм. Допускается, что паразитизм появился вскоре после возникновения жизни. В процессе эволюции пары сожителей могут видоизменяться так, что один партнер начинает как-либо вредить другому. В итоге симбиоз переходит в паразитизм (один из симбиоза становится паразитом, другой - хозяином паразита). Другой путь возникновения паразитизма может быть связан с приспособлением мелких организмов к повторному питанию за счет живого организма хозяина, которому при этом наносится ущерб. Наконец, часть паразитов развилась благодаря способности их предков некоторое время жить в каких-либо хозяевах на положении ложных паразитов. Возникновение паразитизма шло по-разному у экто- и эндопаразитов. Эктопаразитизм формировался прежде всего у свободноживущих хищных клещей, насекомых и других животных за счет удлинения сроков питания и времени пребывания на хозяине. Большую роль в эволюции эктопаразитов сыграли переход от полифагии к монофагии и специализации питания, в частности, питание кровью (клещи, комары, москиты, вши, кровососущие мухи). Эндопаразитизм, в частности, кишечный как наиболее распространенный, формировался в результате случайного заноса цист простейших и яиц гельминтов в желудчно-кишечный тракт животного или человека. В результате случайных контактов у паразитов вырабатывались такие особенности, которые способствовали установлению паразитических отношений. Эндопаразитизм мог возникнуть также в результате изменения инстинкта откладки яиц не на гниющий органический материал, а на раневую поверхность или в полости тела человека, сообщающиеся с внешней средой (вольфартова муха). Кровепаразитизм рассматривается как вторичное явление, явившееся следствием первичного кишечного паразитизма. При этом считают, что современные кровепаразиты позвоночных (например, плазмодии) были кишечными паразитами беспозвоночных, а с переходом хозяев к гематофагии приспособились к жизни в кровяном русле позвоночных, сохранив при этом связь и с беспозвоночными. У одноклеточных в процессе эволюции сформировались органоиды передвижения (у трипаносом); для проникновения в цитоплазму клетки хозяина - коноид. Многоклеточные организмы в процессе адаптации к паразитизму потеряли ряд морфологических структур. Например, уцестод нет кишечника, слабо выражен мышечный слой. Наряду с этим возникли и адаптации прогрессивного характера: мощные органы фиксации (присоски, ботрии, крючья), большая плодовитость, быстрый темп роста, роль кутикулы как защитного барьера от пищеварительных ферментов хозяина и т.д.
Билет 5 В1.Гаметогенез. Строение половых клеток. Гаметогенез подразделяется на сперматогенез (процесс образования сперматозоидов у самцов) и оогенез(процесс образования яйцеклетки). По тому, что происходит с ДНК, эти процессы практически не отличаются: одна исходная диплоидная клетка дает четыре гаплоидные. Однако, по тому, что происходит с цитоплазмой, эти процессы кардинально различаются. В яйцеклетке накапливаются питательные вещества, необходимые в дальнейшем для развития зародыша, поэтому яйцеклетка – это очень крупная клетка, и когда она делится, цель – сохранить питательные вещества для будущего зародыша, поэтому деление цитоплазмы несимметрично. Для того чтобы сохранить все запасы цитоплазмы и при этом избавиться от ненужного генетического материала, от цитоплазмы отделяются полярные тельца, которые содержат очень мало цитоплазмы, но позволяют поделить хромосомный набор. Полярные тельца отделяются при первом и втором делении мейоза
В2.Кариотип человека. Денверская и Парижская Классификация хромосом. Кариоти́п — совокупность признаков (число, размеры, форма и т. д.) полного набора хромосом, присущая клеткам данного биологического вида (видовой кариотип), данного организма (индивидуальный кариотип) или линии (клона) клеток. Кариотипом иногда также называют и визуальное представление полного хромосомного набора (кариограммы). Диверге́нция (от средневекового лат. divergo — отклоняюсь) (в биологии) — расхождение признаков и свойств у первоначально близких групп организмов в ходе эволюции, результат обитания в разных условиях и неодинаково направленного естественного или искусственного отбора
В3.Пути морфо-физиологической адаптации к паразитическому образу жизни. Переход к паразитическому образу жизни сопровождается появлением у паразитов ряда адаптации, облегчающих их существование, развитие и размножение в специфических условиях организма хозяина Разнообразие форм паразитизма, различное систематическое положение паразитов (их принадлежность к разным отрядам, классам и типам), а также обитание их в разных органах и системах хозяина обусловливают многообразие этих адаптации. Однако некоторые приспособления являются абсолютно универсальными. К ним в первую очередь относятся высокая плодовитость и особенности половой системы. Действительно, возможность оставления потомства и попадания его в благоприятную среду — организм хозяина — у паразитов часто ничтожна. В связи с этим интенсивность размножения паразитов по сравнению со свободноживущими формами гораздо более велика. Достигается это разными способами. У многоклеточных это сильная степень развития половой системы и образование огромного количества половых продуктов. Этому способствуют первичный гермафродитизм плоских червей, изначально высокая плодовитость круглых червей и основной массы членистоногих. Нередко высокая интенсивность полового размножения дополняется размножением личиночных стадий жизненного цикла. Особенно это характерно для сосальщиков, личинки которых размножаются партеногенетически, а у некоторых ленточных червей — внутренним или наружным почкованием. Присасывательные диски у инфузории и лямблии, сложная присоска и кутикулярные шипики на коже у сосальщиков, крючья и сложные присоски на головках ленточных червей, конечности, служащие для прикрепления, у ракообразного и насекомого Некоторые паразиты из типа простейших приобретают способность к множественному делению — шизогонии, когда из одного паразита может образоваться более 1000 дочерних особей, или к спорогонии, в результате которой из одной особи могут образоваться десятки тысяч организмов следующего поколения. Практически у всех эктопаразитов и паразитов, обитающих в полостных органах, имеются адаптации для прикрепления к телу хозяина. Они встречаются у простейших (присасывательные диски лямблии), у гельминтов (присоски, шипики, крючья плоских червей, хитинизированный ротовой аппарат ряда круглых червей) и паразитических членистоногих (своеобразные конечности).Эндопаразиты, обитающие в полостных органах, имеют покровы, обладающие антиферментными свойствами, быстро регенерирующие либо вообще непроницаемые для ферментов хозяина. Паразиты, живущие в тканях, часто там инкапсулируются.Паразиты, питающиеся кровью (представлены в основном членистоногими), имеют колюще-сосущий ротовой аппарат, а также сильно растяжимый хитиновый покров, часто разветвленную пищеварительную трубку, антикоагулянтные свойства слюны и консервантные свойства ферментов пищеварительной системы.Эндопаразиты, активно отыскивающие хозяина, обладают органами ориентации в среде, используемыми для поисков хозяина (светочувствительные глазки, термо- и хеморецепторы), и органами передвижения.Передний конец тела паразитов, внедряющихся в организм хозяина, снабжен органами проникновения — специализированными железами, колющими стилетами и т.д. Это касается даже некоторых простейших, способных проникать в ткани хозяина через неповрежденные покровы.Все паразиты, развивающиеся со сменой хозяев, используют в качестве таковых виды, связанные между собой непосредственными пищевыми взаимоотношениями или обитающие с ними в одной среде. Большинство промежуточных хозяев являются источником питания для основных. Другой распространенный путь попадания паразита в организм хозяина — это использование многочисленных переносчиков, которые обеспечивают не только постоянную циркуляцию паразитов в экологических системах, но и их широкое расселение.Высшей степенью адаптации паразитов к хозяевам является наблюдаемая часто полная зависимость паразита от жизнедеятельности хозяев. При этом паразит нередко вызывает такие реакции хозяина, которые обеспечивают максимальную вероятность заражения последнего. Так, самки остриц, откладывая яйца в области анального отверстия, вызывают зуд. Расчесывание зудящих мест способствует распространению яиц этого паразита руками по окружающим предметам. Таким же образом обеспечивается расселение чесоточного клеща. Зуд в пораженной конечности, прекращающийся от соприкосновения с водой, способствует циркуляции в природе такого паразита, как ришта. Высокая температура больных паразитарными заболеваниями, распространяющимися с помощью кровососущих членистоногих, привлекает переносчиков нередко с больших расстояний и также оказывается полезной для паразитов.Нередко особенности жизнедеятельности паразитов оказываются синхронизированными с образом жизни хозяев. Так, откладка яиц шистосомами происходит обычно в самое жаркое время суток, когда наиболее вероятным оказывается контакт хозяев с водой, куда для развития должны попасть яйца этих паразитов. В это же время в поверхностных слоях воды скапливаются в поисках хозяев церкарии этих шистосом. Таким образом облегчается циркуляция паразита сразу на двух стадиях его жизненного цикла. Если в циркуляцию паразита включены несколько хозяев, то наблюдаются их взаимные адаптации, оказывающиеся выгодными паразитам и обеспечивающие его эффективное развитие. Так, выход микрофилярий в кровеносные сосуды человека происходит в часы суток, соответствующие периоду максимальной активности кровососущих насекомых, являющихся их переносчиками.Нередко паразиты даже модифицируют поведение одних хозяев таким образом, что в результате облегчается их попадание к другим. Так, рыбы, пораженные личинками ленточных червей, плавают в основном у поверхности воды и чаще вылавливаются рыбаками и хищными животными. Ленточные черви, использующие в качестве промежуточных хозяев копытных животных, снижают их жизнеспособность, и, таким образом, хищники поедают их в первую очередь. Сосальщики, заражающие травоядных животных и человека через случайное проглатывание насекомых, вызывают обездвиживание последних и плотное прикрепление к растениям, облегчающее им попадание в пищеварительный тракт хозяина.Одновременно с перечисленными признаками свойства паразитов переживать неблагоприятные условия внешней среды являются также несомненными адаптациями к паразитизму. Большинство простейших, заражение которыми происходит без участия переносчиков, во внешней среде способны инцистироваться. Яйца большинства гельминтов обладают феноменальной устойчивостью к неблагоприятным воздействиям. Капсулы с личинками трихинелл переносят не только промораживание, кипячение, но и многократное прохождение через пищеварительную систему рыб, земноводных, птиц, насекомых и ракообразных, не теряя жизнеспособности.Для большинства паразитов, переживших нахождение во внешней среде или в промежуточном хозяине в покоящихся стадиях и попавших в организм окончательного хозяина, существует комплекс условий, являющийся сигналом к началу активной жизнедеятельности. У млекопитающих он часто неспецифичен: это температура тела около 37°С, водная среда и высокая ее кислотность, а также наличие ферментов желудочного сока. Такие условия характерны для желудка любого млекопитающего, поэтому цисты, яйца и другие инвазионные стадии паразитов, обитающие у разных млекопитающих могут, попадая в желудок человека, начинать развитие. Некоторые из них проходят лишь часть цикла и, не находя специфических условий, гибнут успевая, однако, привести к тяжелым последствиям.Указанные особенности паразитов, общие для многих из них, не состоящих в родстве, возникли в разных их группах независимо друг от друга, иллюстрируя конвергентный характер эволюции организмов разных видов, классов и типов, адаптирующихся к сходным условиям.
Билет 6 В1.Цитологические основы полового размножения. Мейоз, как специфический процесс при формировании половых клеток. Мейоз (от греч.meiosis — уменьшение) или редукционное деление клетки — деление ядра эукариотическойклетки с уменьшением числа хромосом в два раза. Происходит в два этапа (редукционный и эквационный этапы мейоза). Мейоз не следует смешивать с гаметогенезом — образованием специализированных половых клеток, или гамет, из недифференцированныхстволовых. С уменьшением числа хромосом в результате мейоза в жизненном цикле происходит переход от диплоидной фазы к гаплоидной. Восстановление плоидности (переход от гаплоидной фазы к диплоидной) происходит в результате полового процесса. В связи с тем, что в профазе первого, редукционного, этапа происходит попарное слияние (конъюгация) гомологичных хромосом, правильное протекание мейоза возможно только в диплоидных клетках или в чётных полиплоидах (тетра-, гексаплоидных и т. п. клетках). Мейоз может происходить и в нечётных полиплоидах (три-, пентаплоидных и т. п. клетках), но в них, из-за невозможности обеспечить попарное слияние хромосом в профазе I, расхождение хромосом происходит с нарушениями, которые ставят под угрозу жизнеспособность клетки или развивающегося из неё многоклеточного гаплоидного организма. Этот же механизм лежит в основе стерильности межвидовых гибридов. Поскольку у межвидовых гибридов в ядре клеток сочетаются хромосомы родителей, относящихся к различным видам, хромосомы обычно не могут вступить в конъюгацию. Это приводит к нарушениям в расхождении хромосом при мейозе и, в конечном счете, к нежизнеспособности половых клеток, или гамет. Определенные ограничения на конъюгацию хромосом накладывают и хромосомные мутации (масштабные делеции, дупликации, инверсии или транслокации). Мейоз состоит из 2 последовательных делений с короткой интерфазой между ними.
· Лептотена или лептонема — упаковка хромосом. · Зиготена или зигонема — конъюгация (соединение) гомологичных хромосом с образованием структур, состоящих из двух соединённых хромосом, называемых тетрадами или бивалентами. · Пахитена или пахинема — кроссинговер (перекрест), обмен участками между гомологичными хромосомами; гомологичные хромосомы остаются соединенными между собой. · Диплотена или диплонема — происходит частичная деконденсация хромосом, при этом часть генома может работать, происходят процессы транскрипции (образование РНК), трансляции (синтез белка); гомологичные хромосомы остаются соединёнными между собой. У некоторых животных в ооцитах хромосомы на этой стадии профазы мейоза приобретают характерную форму хромосом типа ламповых щёток. · Диакинез — ДНК снова максимально конденсируется, синтетические процессы прекращаются, растворяется ядерная оболочка; центриоли расходятся к полюсам; гомологичные хромосомы остаются соединёнными между собой.
Второе деление мейоза следует непосредственно за первым, без выраженной интерфазы: S-период отсутствует, поскольку перед вторым делением не происходит репликации ДНК.
В результате из одной диплоидной клетки образуется четыре гаплоидных клетки. В тех случаях, когда мейоз сопряжён с гаметогенезом (например, у многоклеточных животных), при развитии яйцеклеток первое и второе деления мейоза резко неравномерны. В результате формируется одна гаплоидная яйцеклетка и два так называемых редукционных тельца (абортивные дериваты первого и второго делений).
В2.Методы в генетике человека. Цитогенетические методы. Кариотип человека. Генеалогический метод, Популяционный метод., Близнецовый метод. Цитологический метод, Биохимический метод. Цитогенетические методы исследования применяют для диагностики хромосомных болезней. Они включают:
Определение Х- и Y-хроматина Определение Х- и Y-хроматина часто называют методом экспресс-диагностики пола. Исследуют клетки слизистой оболочки ротовой полости вагинального эпителия или волосяной луковицы. В ядрах клеток женщин в диплоидном наборе присутствуют две хромосомы X, одна из которых полностью инактивирована (спирализована, плотно упакована) уже на ранних этапах эмбрионального развития и видна в виде глыбки гетерохроматина, прикреплённого к оболочке ядра. Инактивированная хромосома X называется половым хроматином или тельцем Барра. Для выявления полового Х-хроматина (тельца Барра) в ядрах клеток мазки окрашивают ацетарсеином и препараты просматривают с помощью обычного светового микроскопа. В норме у женщин обнаруживают одну глыбку Х-хроматина, а у мужчин её нет. Для выявления мужского Y-полового хроматина (F-тельце) мазки окрашивают акрихином и просматривают с помощью люминисцентного микроскопа. Y-хроматин выявляют в виде сильно светящейся точки, по величине и интенсивности свечения отличающейся от остальных хромоцентров. Он обнаруживается в ядрах клеток мужского организма. Отсутствие тельца Барра у женщин свидетельствует о хромосомном заболевании - синдроме Шерешевского-Тернера (кариотип 45, ХО). Присутствие у мужчин тельца Барра свидетельствует о синдроме Кляйнфелтера (кариотип 47, XXY). Определение Х- и Y-хроматина - скрининговый метод, окончательный диагноз хромосомной болезни ставят только после исследования кариотипа. Кариотипирование Для изучения хромосом чаще всего используют препараты кратковременной культуры крови, а также клетки костного мозга и культуры фибробластов. Доставленную в лабораторию кровь с антикоагулянтом подвергают центрифугированию для осаждения эритроцитов, а лейкоциты инкубируют в культуральной среде 2-3 дня. К образцу крови добавляют фитогемагглютинин, так как он ускоряет агглютинацию эритроцитов и стимулирует деление лимфоцитов. Наиболее подходящая фаза для исследования хромосом - метафаза митоза, поэтому для остановки деления лимфоцитов на этой стадии используют колхицин. Добавление этого препарата к культуре приводит к увеличению доли клеток, находящихся в метафазе, то есть в той стадии клеточного цикла, когда хромосомы видны лучше всего. Каждая хромосома реплицируется (производит свою копию) и после соответствующей окраски видна в виде двух хроматид, прикреплённых к центромере, или центральной перетяжке. Затем клетки обрабатывают гипотоническим раствором хлорида натрия, фиксируют и окрашивают. Для окраски хромосом чаще используют краситель Романовского-Гимзы, 2% ацеткармин или 2% ацетарсеин. Они окрашивают хромосомы целиком, равномерно (рутинный метод) и могут быть использованы для выявления численных аномалий хромосом человека. Для получения детальной картины структуры хромосом, идентификации (определения) отдельных хромосом или их сегментов используют различные способы дифференциального окрашивания. Наиболее часто применяют методы Гимза, а также G- и Q-бендинга. При микроскопии препарата по длине хромосомы выявляется ряд окрашенных (гетерохроматин) и неокрашенных (эухроматин) полос. Характер поперечной исчерченности, получаемый при этом, позволяет идентифицировать каждую хромосому в наборе, так как чередование полос и их размеры строго индивидуальны и постоянны для каждой пары.Метафазные пластинки отдельных клеток фотографируют. Из фотографий вырезают индивидуальные хромосомы и наклеивают их по порядку на лист бумаги; такая картина хромосом называется кариотипом.Применение дополнительного окрашивания, а также новые методы получения хромосомных препаратов, позволяющих растягивать хромосомы в длину, значительно увеличивают точность цитогенетической диагностики.Для описания кариотипа человека разработана специальная номенклатура. Нормальный кариотип мужчины и женщины обозначают как 46, XY и 46, XX соответственно. При синдроме Дауна, характеризующемся наличием дополнительной хромосомы 21 (трисомия 21), кариотип женщины описывают как 47, XX 21+, а мужчины - 47, XY, 21+. При наличии структурной аномалии хромосомы необходимо указать изменённое длинное или короткое плечо: буквой р обозначают короткое плечо, q - длинное плечо, t - транслокацию. Так, при делеции короткого плеча хромосомы 5 (синдром «кошачьего крика») женский кариотип описывают как 46, XX, 5р-. Мать ребёнка с транслокационным синдромом Дауна - носительница сбалансированной транслокации 14/21 имеет кариотип 45, XX, t(14q; 21q). Транслокационная хромосома образуется при слиянии длинных плеч хромосомы 14 и 21, короткие плечи при этом теряются. Каждое плечо разделяется на районы, а они в свою очередь - на сегменты, и те и другие обозначают арабскими цифрами. Центромера хромосомы является исходным пунктом для отсчёта районов и сегментов. Таким образом, для топографии хромосом используют четыре метки: номер хромосомы, символ плеча, номер района и номер сегмента в пределах данного района. Например, запись 6р21.3 означает, что речь идёт о хромосоме 6-й пары, её коротком плече, районе 21, сегменте 3. Существуют ещё дополнительные символы, в частности pter - конец короткого плеча, qter - конец длинного плеча. Цитогенетический метод исследования позволяет обнаружить делеции и другие изменения в хромосомах только размером приблизительно в 1 млн оснований (нуклеотидов).
В3. Понятие об трансмиссивных болезнях. Экологические основы их выведения. Трансмиссивные болезни (лат. transmissio —перенесение на других) — заразные болезни человека, возбудители которых передаются кровососущими членистоногими (насекомыми и клещами). Трансмиссивные болезни включают более 200 нозологических форм, вызываемых вирусами, бактериями, риккетсиями, простейшими и гельминтами. Часть из них передаётся только с помощью кровососущих переносчиков (облигатные трансмиссивные болезни, например сыпной тиф, малярия и др.), часть различными способами, в том числе и трансмиссивно (например, туляремия, заражение которой происходит при укусах комаров и клещей, а также при снятии шкурок с больных животных). В передаче возбудителей трансмиссивных болезней участвуют специфические и механические переносчики. В специфических переносчиках возбудитель либо размножается и накапливается (например, вирусы в теле клещей), либо созревает до инвазионной стадии (например, личинки филярий в комарах, мошках, слепнях, малярийные плазмодии в теле комаров). Как размножение, так и развитие возбудителей в теле специфических переносчиков может происходить только при достаточно высокой, строго определённой температуре для каждой пары паразит — переносчик. Так, половое размножение, или спорогония, возбудителя трёхдневной малярии продолжается 19 дней при 20 °C и 6½ дней при температуре 30 °C, и только после этого в слюнных железах переносчика появляется инвазионная стадия паразита — спорозоит. Передача возбудителя переносчиком осуществляется различными способами. Наиболее часто возбудитель проникает со слюной, которую переносчик вводит в место укола перед началом кровососания. Так вводятся спорозоиты малярийного паразита. Перенос риккетсий вшами (см. Сыпной тиф эпидемический) и трипаносом триатомовыми клопами происходит во время дефекации переносчика; возбудитель из фекалий проникает затем в ранку на месте укола. Возбудитель вшиного возвратного тифа (Borrelia recurrentis) скапливается в теле вши и может проникнуть в организм человека лишь при раздавливании вши руками и втирании ее гемолифмы в кожу при расчёсах. Многие переносчики передают лишь одного, специфического для них паразита. Так, мухи цеце передают лишь трипаносому — возбудителя сонной болезни человека, мошки — возбудителя онхоцеркоза, триатомовые клопы — возбудителя американского трипаносомоза (болезни Шагаса). Москиты могут переносить возбудителей лейшманиозов и лихорадки паппатачи. Наиболее разнообразные связи с паразитами установились у комаров и клещей. Комары семейства Culicidae передают многие виды вирусов, в том числе возбудителя жёлтой лихорадки, лихорадки денге, вирусных энцефалитов (см. Энцефалиты комариные). Они являются переносчиками также малярии и филяриатозов. Иксодовые клещи передают вирусы, в том числе возбудителей клещевого энцефалита и геморрагических лихорадок, риккетсий — возбудителей пятнистых лихорадок, североазиатского сыпного тифа (см. Североазиатский клещевой риккетсиоз) и др., а также спирохет. В переносе возбудителей болезни у многих групп насекомых участвуют только самки. У мухи цеце, вшей и у всех групп клещей кровососами и переносчиками являются особи обоего пола. В ряде случаев возбудитель болезни передается переносчиком своему потомству. Такая трансовариальная передача выявлена у комаров, инфицированных вирусами, у клещей, инфицированных вирусами и спирохетами и у москитов, инфицированных флебовирусами. В организме механических переносчиков возбудители не развиваются и не размножаются. Попавший на хоботок, в кишечник или на поверхность тела механического переносчика возбудитель передается непосредственно (при укусе) либо путем контаминации ран, слизистых оболочек хозяина или пищевых продуктов. Самыми распространенными механическими переносчиками являются мухи семейства Muscidae, которые известны как переносчики вирусов, бактерий, простейших, гельминтов. Билет 7 В1. Принцип компартмации. Биологическая мембрана.
В2.Ген – функциональная единица наследственности. Эволюция представлений о гене. Ген — структурная и функциональная единица наследственности, контролирующая развитие определенного признака или свойства. Совокупность генов родители передают потомкам во время размножения. В настоящее время, в молекулярной биологии установлено, что гены — это участки ДНК, несущие какую-либо целостную информацию — о строении одной молекулыбелка или одной молекулы РНК. Эти и другие функциональные молекулы определяют развитие, рост и функционирование организма. В то же время, каждый ген характеризуется рядом специфических регуляторных последовательностей ДНК, таких как промоторы, которые принимают непосредственное участие в регулировании проявления гена. Регуляторные последовательности могут находиться как в непосредственной близости от открытой рамки считывания, кодирующей белок, или начала последовательности РНК, как в случае с промоторами (так называемые cis-регуляторные элементы, англ. cis-regulatory elements), так и на расстоянии многих миллионов пар оснований (нуклеотидов), как в случае с энхансерами, инсуляторами и супрессорами (иногда классифицируемые как trans-регуляторные элементы, англ. trans-regulatory elements). Таким образом, понятие гена не ограничено только кодирующим участком ДНК, а представляет собой более широкую концепцию, включающую в себя и регуляторные последовательности. Изначально термин ген появился как теоретическая единица передачи дискретной наследственной информации. История биологии помнит споры о том, какие молекулы могут являться носителями наследственной информации. Большинство исследователей считали, что такими носителями могут быть только белки, так как их строение (20 аминокислот) позволяет создать больше вариантов, чем строение ДНК, которое составлено всего из четырёх видов нуклеотидов. Позже было экспериментально доказано, что именно ДНК включает в себя наследственную информацию, что было выражено в виде центральной догмы молекулярной биологии. Гены могут подвергаться мутациям — случайным или целенаправленным изменениям последовательности нуклеотидов в цепи ДНК. Мутации могут приводить к изменению последовательности, а следовательно изменению биологических характеристик белка или РНК, которые, в свою очередь, могут иметь результатом общее или локальное изменённое или анормальное функционирование организма. Такие мутации в ряде случаев являются патогенными, так как их результатом является заболевание, или летальными на эмбриональном уровне. Однако, далеко не все изменения последовательности нуклеотидов приводят к изменению структуры белка (благодаря эффекту вырожденности генетического кода) или к существенному изменению последовательности и не являются патогенными. В частности, геном человека характеризуется однонуклеотидными полиморфизмами и вариациями числа копий (англ. copy number variations), такими как делеции и дупликации, которые составляют около 1 % всей нуклеотидной последовательности человека.[1] Однонуклеотидные полиморфизмы, в частности, определяют различные аллели одного гена. Мономеры, составляющие каждую из цепей ДНК, представляют собой сложные органические соединения, включающие в себя азотистые основания: аденин(А) или тимин(Т) или цитозин(Ц) или гуанин(Г), пятиатомный сахар-пентозу-дезоксирибозу,по имени которой и получила название сама ДНК, а так же остаток фосфорной кислоты.Эти соединения носят название нуклеотидов. |
||||
|
|
Последнее изменение этой страницы: 2018-04-12; просмотров: 559. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |

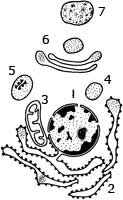 Компартментация объема клетки с помощью мембран
Высокая упорядоченность внутреннего содержимого эукариотической клетки достигается путем компартментации ее объема — подразделения на «ячейки», отличающиеся деталями химического (ферментного) состава. Компартментация способствует пространственному разделению веществ и процессов в клетке. Отдельный компартмент представлен органеллой (лизосома) или ее частью (пространство, отграниченное внутренней мембраной митохондрии).
1—ядро, 2—шероховатая цитоплазматическая есть, 3—митохондрия, 4 - транспортный цитоплазматический пузырек, 5—лизосома, 6—пластинчатый комплекс, 7 —гранула секрета. Предложено несколько схем взаимоотношения вмембране основных химических компонентов - белков и липидов, а также веществ, размещаемых на мембранной поверхности. В настоящее время принята точка зрения, согласно которой мембрана составлена из бимолекулярного слоя липидов. Гидрофобные участки их молекул повернуты друг к другу, а гидрофильные - находятся на поверхности слоя. Разнообразные белковые молекулы встроены в этот слой или размещены на его поверхностях. Благодаря компартментации клеточного объема в эукариотической клетке наблюдается разделение функций между разными структурами. Одновременно различные структуры закономерно взаимодействуют друг с другом.
Компартментация объема клетки с помощью мембран
Высокая упорядоченность внутреннего содержимого эукариотической клетки достигается путем компартментации ее объема — подразделения на «ячейки», отличающиеся деталями химического (ферментного) состава. Компартментация способствует пространственному разделению веществ и процессов в клетке. Отдельный компартмент представлен органеллой (лизосома) или ее частью (пространство, отграниченное внутренней мембраной митохондрии).
1—ядро, 2—шероховатая цитоплазматическая есть, 3—митохондрия, 4 - транспортный цитоплазматический пузырек, 5—лизосома, 6—пластинчатый комплекс, 7 —гранула секрета. Предложено несколько схем взаимоотношения вмембране основных химических компонентов - белков и липидов, а также веществ, размещаемых на мембранной поверхности. В настоящее время принята точка зрения, согласно которой мембрана составлена из бимолекулярного слоя липидов. Гидрофобные участки их молекул повернуты друг к другу, а гидрофильные - находятся на поверхности слоя. Разнообразные белковые молекулы встроены в этот слой или размещены на его поверхностях. Благодаря компартментации клеточного объема в эукариотической клетке наблюдается разделение функций между разными структурами. Одновременно различные структуры закономерно взаимодействуют друг с другом.