|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
Влияние исследуемых антидепрессантов на эффекты дексаметазона в исследованиях на нейронах дорсального стриатума.Объект исследования синаптическая передача между аксонами пирамидных нейронов сомато-сенсорной коры, проходящих в мозолистом теле, и аксонами среднешипиковых ГАМК- ергических нейронов дорсального стриатума.
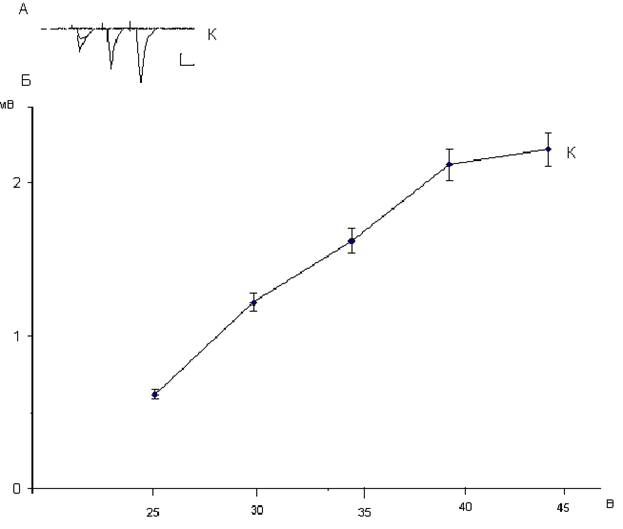
Рис. 1.13. Зависимость амплитуд популяционных спайков нейронов дорсального стриатума от интенсивности стимуляции кортико-стриатных волокон. Фокальные потенциалы, генерируемые исследуемыми нейронами (А), и зависимость их амплитуды от интенсивности пресинаптической стимуляции (Б). На А: калибровка 1 мВ и 10 мс. На Б: вертикальная шкала – амплитуда потенциалов в мВ, горизонтальная шкала – интенсивность пресинаптической стимуляции в В. В связи с особенностями архитенктоники дорсо - медиального стриатума популяционные ВПСП среднешипиковых проекционных нейронов в условиях внеклеточной микроэлектродной регистрации имеют небольшую амплитуду, потому мы регистрировали популяционные спайки этих нейронов, которые являются ортодромными, а не антидромными фокальными потенциалами. Действительно, поскольку регистрируемые популяционные спайки нейронов дорсального стриатума обратимо угнетается при нарушении процессов везикулярного высвобождения глутамата при блокаде потенциалозависимых кальциевых каналов в пресинаптических аксонных терминалях ионами магния и кадмия (не иллюстрировано), они имеют постсинаптическую природу и генерируются на вершинах ВПСП. По сравнению с постсинаптическими ответами пирамидных и зернистых нейронов гиппокампа, для генерации постсинаптических ответов дорсального стриатума требуется более мощная пресинаптическая стимуляция (рис. 1.1, 1.7 и 1.13). Это обусловлено двумя моментами. Во-первых, среднешипиковые нейроны дорсального стриатума имеют самый высокий мембранный потенциал > -80 мВ [131]. Во-вторых, волокна кортикостриатных нейронов имеют выраженную миелиновую оболочку и плотно упакованы среди других миелинизированных аксонов в мозолистом теле. Поэтому для того, чтобы вызвать популяционные спайки среднешипиковых нейронов дорсального стриатума требуется достаточно интенсивная стимуляция мозолистого тела.  Через 24 часа после внутрибрюшинного введения крысам дексаметазона в дозе 7 мг/кг амплитуда популяционных спайков нейронов дорсального стриатума претерпела изменения в сравнении с постсинаптическими ответами в контроле при тех же интенсивностях пресинаптической стимуляции (рис. 1.14).
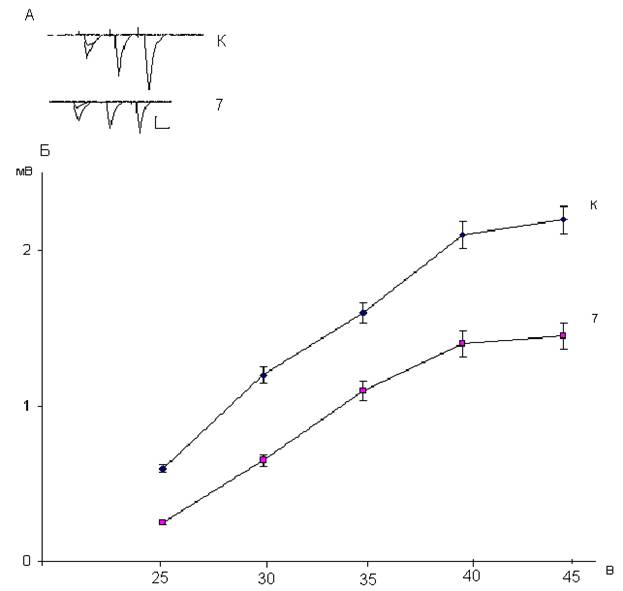
Рис. 1.14. Зависимость амплитуд популяционных спайков нейронов дорсального стриатума от интенсивности пресинаптической стимуляции у контрольных (К) и подвергнутых воздействию дексаметазона в дозе 7 мг/ кг крыс (7). Фокальные потенциалы, генерируемые исследуемыми нейронами (А), и зависимость их амплитуды от интенсивности пресинаптической стимуляции (Б). На А: калибровка 1 мВ и 10 мс. На Б: вертикальная шкала – амплитуда потенциалов в мВ, горизонтальная шкала – интенсивность пресинаптической стимуляции в В.
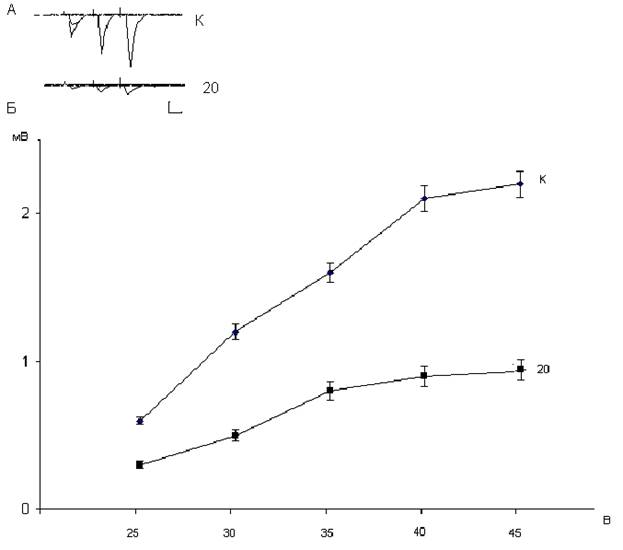
Рис. 1.15. Зависимость амплитуд популяционных спайков нейронов дорсального стриатума от интенсивности пресинаптической стимуляции у контрольных (К) и подвергнутых воздействию дексаметазона в дозе 20 мг/кг крыс (20). Фокальные потенциалы, генерируемые исследуемыми нейронами (А), и зависимость их амплитуды от интенсивности пресинаптической стимуляции (Б). На А: калибровка 1 мВ и 10 мс. На Б: вертикальная шкала – амплитуда потенциалов в мВ, горизонтальная шкала – интенсивность пресинаптической стимуляции в В. Нейротоксическое действие дексаметазона, используемого в дозе 7 мг/кг проявляется снижением амплитуд популяционных спайков нейронов дорсального стриатума на 30-50% и в меньшей степени снижением наклона кривой зависимости амплитуд постсинаптических ответов от интенсивности пресинаптической стимуляции (рис.1.14). При введении крысам большей (20 мг/кг) дозы дексаметазона нейрон-повреждающее действие стероида усиливалось. Это проявлялось более значительным снижением амплитуд популяционных спайков нейронов дорсального стриатума – на 50-70% и более существенным снижением наклона кривой, характеризующей зависимость величин постсинаптических ответов среднешипиковых нейронов от интенсивности пресинаптической стимуляции (рис. 1.15). Для выяснения локализации вызываемого дексаметазоном повреждения нейропередачи в кортико-стриатных синапсах мы исследовали изменения величин парного облегчения популяционных спайков нейронов дорсального стриатума при межимпульсном интервале 50 мс. Таблица 1.3. Величины парного облегчения в % популяционных спайков нейронов дорсального стриатума.
Как следует из таблицы 1.3 величины парного облегчения популяционных спайков нейронов дорсального стриатума при межимпульсном интервале 50 мс у контрольных и подвергнутых воздействию дексаметазона в дозах 7 и 20 мг/кг не претерпевают существенных изменений. Этот факт делает приемлемой гипотезу, согласно которой дексаметазон вызывает повреждение постсинаптических уплотнений в дендритных шипиках нейронов дорсального стриатума. В дальнейшем эффекты дексаметазона можно охарактеризовать по уменьшению синаптической реактивности, которая определяется отношением амплитуд постсинаптических ответов в мВ к интенсивности стимуляции в В. Вызываемые используемыми дозами дексаметазона изменения синаптической реактивности представлены в следующем рисунке. Как видно из рис. 1.16 изменения величин синаптической реактивности среднешипиковых нейронов дорсального стриатума в определенной степени параллельны изменениям кривых зависимости величин популяционных спайков от интенсивности пресинаптической стимуляции (рис. 1.13-1.15). Однако характеристика функционального состояния среднешипиковых нейронов дорсального стриатума по критерию синаптической реактивности более рельефна и более наглядна по сравнению с кривыми зависимости величины постсинаптического ответа от интенсивности пресинаптической стимуляции.
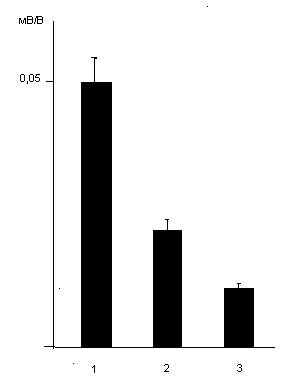
Рис. 1.16. Изменения синаптической реактивности среднешипиковых нейронов дорсального стриатума, вызываемые используемыми дозами дексаметазона. 1- контроль; 2 и 3 – через 24 часа после введения дексаметазона в дозах 7 и 20 мг/кг соответственно. По вертикальной шкале – синаптическая реактивность в мВ/В.
Для уточнения механизмов нейронповреждающего действия дексаметазона мы провели нейрохимический анализ. Исследовали влияние веществ – анализаторов на вызываемое дексаметазоном, используемым в дозе 7 мг/кг, на синаптическую реактивность нейронов дорсального стриатума.
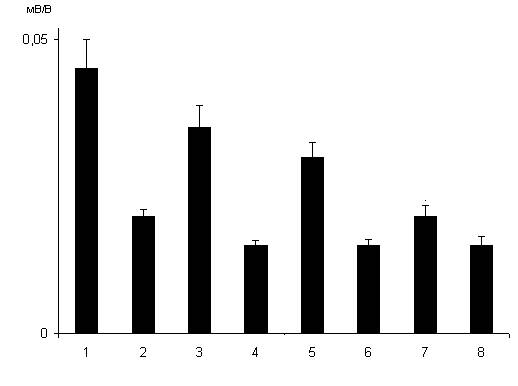
Рис. 1.17. Влияние веществ-анализаторов на вызываемое дексаметазоном (7 мг/кг) угнетение синаптической реактивности нейронов дорсального стриатума. 1 – контроль; 2 – через 24 часа после введения 7 мг/кг дексаметазона; 3 – 8 – то же в условиях введения крысам кетамина 50 мг/кг (3), глицина 50 мг/кг (4), натрия ортованадата 15 мг/кг (5), имипрамина 20 мг/кг (6), флуоксетина 20 мг/кг (8) и пиразидола 20 мг/кг (7).
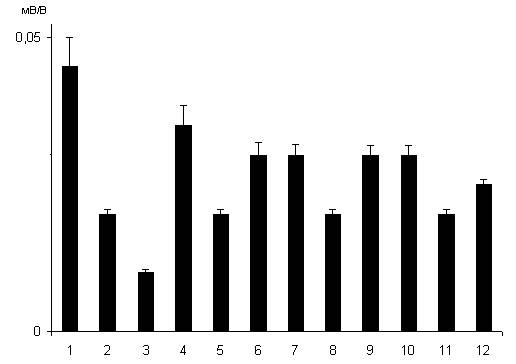
Рис. 1.18. Влияние хронического введения исследуемых антидепрессантов на вызываемое дексаметазоном угнетение синаптической реактивности нейронов дорсального стриатума. 1 – контроль; 2 и 3 – через 24 часа после введения дексаметазона в дозах 7 и 20 мг/кг соответственно; 4,7 и 10 – эффекты дексаметазона (7 мг/кг) после хронического введения имипрамина, флуоксетина и пиразидола; 5, 8 и 11 – эффекты дексаметазона (20 мг/кг) после хронического введения имипрамина, флуоксетина и пиразидола; 6, 9 и 12 – то же, что и на 5, 8 и 11 на фоне предварительного введения 15 мг/кг натрия ортованадата.
Вызываемое дексаметазоном, используемым в дозе 7 мг/кг, угнетение синаптической реактивности проекционных нейронов дорсального стриатума ослаблялось неконкурентным блокатором НМДА рецепторов кетамином (рис. 1.17, столбцы 2 и 3). Однако ко-агонист НМДА глутаматных рецепторов глицин в дозе 50 мг/кг усиливал вызываемое дексаметазоном угнетения синаптической реактивности нейронов дорсального стриатума (рис. 1.17, столбцы 2 и 4). Воздействие ингибитора тирозиновых протеинфосфатаз натрия ортованадата в дозе 15 мг/кг ослабляло вызываемое дексаметазоном угнетение синаптической реактивности стриатных нейронов (рис. 1.17, столбцы 2 и 5). Вводимые однократно в дозе 20 мг/кг антидепрессанты имипрамин и флуоксетин усиливали нейрон-повреждающее действие используемого в дозе 7 мг/кг дексаметазона (рис. 1.17, столбцы 2 и 6,8). Однако антидепрессант из группы ингибиторов МАО А пиразидол, вводимый в дозе 20 мг/кг однократно, не оказывал влияния на эффекты дексаметазона (рис. 1.17, столбцы 2 и 7). Природа нейрон-повреждающего действия дексаметазона в отношении нейронов стриатума в общих чертах сходна с таковой дексаметазона в отношение нейронов гиппокампа. При хроническом введении используемые антидепрессанты в отличие от однократного введения оказывали иное влияние на эффекты дексаметазона. Действительно, все исследуемые антидепрессанты ослабляли угнетение синаптической реактивности среднешипиковых нейронов дорсального стриатума, вызываемое дексаметазоном в дозе 7 мг/кг (рис. 1.18, столбцы 2 и 4,7,9), а также в дозе 20 мг/кг (рис. 1.18, столбцы 3 и 5,8,11). Нейропротективное действие исследуемых антидепрессантов в условиях действия дексаметазона в дозе 20 мг/кг усиливалось натрием ортованадатом (рис. 1.18, столбцы 3 и 6,9,12).
|
||||||||||||||
|
|
Последнее изменение этой страницы: 2018-04-12; просмотров: 295. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |