|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
Regulation activity of genes, sign.6.4.2. Прокариоттар гендерінің белсенділігінің транскрипциялық деңгейдегі реттелуі 1) негативті 2) позитивті 6.4.3. Прокариоттар гендерінің белсенділігінің реттелуінің оперондық моделі 1) индуцибельді оперон 2) репрессибельді оперон 6.4.4. Прокариоттар гендерінің белсенділігінің трансляциялық деңгейдегі реттелуі 6.4.5. Прокариоттар гендерінің белсенділігінің реттелуінің посттрансляциялық деңгейдегі реттелулері * Ағылшын және орыс тілдерінде талдау үшін
Ақпараттық-дидактикалық блок Гендер өнімінің синтезін бақылау механизмдері жалпы атпен гендік реттелу деп аталады. Прокариоттар геномы сақиналы ДНҚ молекуласы нуклеоид және бірнеше майда сақиналы ДНҚ-плазмидалар түрінде болады. Олар үлкен үнемділігімен ерекшеленеді, өйткені онда ақпараты жоқ нуклеотидтер қатары болмайды. Прокариот жасушаларында транскрипция және трансляция үдерісітері қатар жүреді. Геннің өнімі болып ақуыз молекуласының биосинтезі жайлы ақпараты бар а-РНҚ болып табылады. Ақуыздар (полипептидтер) құрылымдық ақуыздар, ферменттер және реттеуші ақуыздар болып бөлінеді. Құрылымдық ақуыздар жасушалармен мүшелердің негізгі құрылым бірлігі. Ферменттер ағззадағы зат алмасуды қамтамасыз етеді. Реттеуші ақуыздар құрылымдық гендердің белсенділігіне әсер етеді: биосинтезді күшейтеді, төмендетеді немесе басып тастайды. Прокариоттарда (ішек таяқшасы E.Coli) кейбір ферменттер сыртқы орта жағдайына тәуелсіз үнемі үздіксіз синтезделеді.  Ағзада конститутивті (тұрақты) ферменттердің синтезделуі, сол ферменттің синтезін бақылайтын геннің үздіксіз белсенді болатынын көрсетеді. Сонымен бірге бірқатар ферменттер ағзаға қажетті жағдайда (ортада субстраттар болғанда) ғана синтезделеді. Егер жасушаға (ағзаға) ферменттер қажет болмайтын болса, онда олардың синтезделуі яғни ферменттердің активтілігін бақылайтын гендердің де жұмыстары тоқтайды. Прокариот гендерінің активтілігінің реттелуі үш деңгейде реттеледі: 1.Транскрипциялық 2. Трансляциялық 3. Посттрансляциялық Прокариоттардың тіршілігін қамтамасыз етуде гендер активтілігінің реттелуінің маңызы зор. Прокариот гендерінің активтілігінің реттелуі: Транскрипциялық бақылаудың екі түрі бар: негативті және позитивті.
Ген активтілігін негативті бақылау жасушада геннің транскрипциясын тоқтататын арнайы факторлар – ингибиторлардың болуына негізделген. Жасушада сонымен қатар транскрипцияның басталуына мүмкіндік беріп, ингибиторларға қарсы әрекет көрсететін индуктор-факторлары болады. Ген активтілігін позитивті бақылау, промоторды активтендіріп, транскрипцияны бастайтын эффекторлық ақуыз молекулалары мен геннің әрекеттесуіне байланысты. Көпшілік жағдайда бақылаудың осы екі түрі де қатарынан жүреді. Мысалы, катаболистік (ыдыратушы) жүйелер позитивтік жолмен де, негативтік жолмен де реттелуі мүмкін. Негативтік бақылау Индукцияланатын жүйелер (индукцияланатын оперондар). Бұл жүйелерде ферменттің синтезі үшін жасушаны қоршаған ортада индуктор деп аталатын арнайы субстрат-затының болуы қажет. Алғаш рет Ф. Жакоб және Ж. Моно (1961) индуцибельді жүйені тәжірибе жүзінде зерттеу негізінде ішек таяқшасындағы (Е.coli)β – галактозидаза ферментінің синтезінің реттелуін Lac – оперон моделін ұсыну арқылы сол ферменттердің синтезделуінің индуцибельді (қоздырушы) немесе репрессибельді (басылыңқы) механизмдерін түсіндірді. Жакоб және Моно бойынша Lac – оперонының құрылысы мен қызметі Лактозалық оперонныі сызбасы
Оперон ақуыз синтезінің бақылануының структуралық- функционалдық бірлігінің элементтері: 1. Құрылымдық гендер - ферменттер синтезіне қатысады 2. Оперон - белок-репрессоры байланысатын сайт 3. Промотор - РНК-полимераза байланысатын сайт Оперон құрамындағы қатарласа орналасқан құрылымдық гендер - кластерлық гендер деп аталады. Осы кластерлық гендерден тұтас бір ғана полицистронды РНҚ түзіледі. Полицистронды РНҚ-дан трансляция нәтижесінде жалпы қызмет атқаратын бірнеше белок-ферменттер синтезделеді. Лактозалық оперондағы (Lac – оперон) құрылымдық гендер үш цистроннан тұрады: Z,Y,A. Бұл гендердің түзілетін өнімі – (ферменттер) лактоза қантын ыдыратуға қатынасады, β-галактозидаза ферментін синтездейтін Z – гені лактозаны глюкоза мен галактозаға дейін ыдырады. Y генінің өнімі – фермент галактозид-пермеаза. Бұл ферменттің қызметі жасушаның мембранасымен байланысып лактозаның жасушаға сіңірілуін қамтамасыз етеді. А-генің өнімі – тигалактозид – ацетилаза ферменті лактозаның толық ыдырауына қатынасады. Қалыпты жағдайда, яғни жасушада лактоза жоқ кезде бұл үш ферменттердің синтезі жүрмейді, осы аталған ферменттердің аздаған молекулалары ғана болуы мүмкін. Жасушаға лактоза сіңірілген кезде барлық үш ферменттің синтезі қарқынды жүре бастайды. Олай болса, лактозаның жасушаға енуі оны ыдыратып пайдалануға қажетті ферменттердің синтезін индукциялайды. Барлық үш фермент бір ғана полицистрондық а-РНҚ-ның трансляциясына байланысты түзілгендіктен бұл жағдайды жасушаға лактоза – индуктордың енуіне жауап ретінде оперонның біртұтас үйлесімді қызметі деп түсіндіреді. Жакоб және Моно теориясы бойынша реттеуші ген (i) репрессор-ақуызын синтездейді, ортада индуктор (лактоза) жоқ кезде ол оператормен (о) байланысып оны активсіздендіреді, Lac –оперондағы үш Lac –цистрондар транскрипцияланбайды. Құрылымдық гендердің транскрипциясының индукциясын (басталуы) былай түсіндіруге болады, жасушаға енген лактоза индуктор ретінде репрессор белогымен байланысып оны оператордан ажыратып активсіздендіреді. Репрессордан ажыраған оператор оперонның жұмыс істеуіне мүмкіндік береді: құрылымдық гендердің үйлесімді транскрипциясы мен трансляциясы басталады. Репрессор белогында екі түрлі қайта жабылмайтын сайттар болады: 1) оператормен байланыстыратын сайт; 2) индуктормен байланысу сайты. Жакоб және Мононың зерттеулері кезінде (1961 жылдары) промотордың бар екендігі және репрессорлық реттелудің механизмі жайлы деректер болмаған. Кейінірек E. coli-дің ДНҚ тізбегінде РНҚ полимераза ферменті жалғанатын промоторлық сайттың бар екндігі және ол сайт репрессор (і) гені мен оператор (о) гендерінің арасында орналасқаны белгілі болды. Репрессор гені Lac – оперонынан бөлек моноцистронды а-РНҚ транскрипциаланып репрессор ақуызын синтездейді. Ағзаға түскен индукторға жауап ретінде барлық үш фермент бір ғана полицистронды а-РНҚ трансляциялануынан синтезделетін болғандықтан оперон бір тұтас жүйе ретінде жұмыс істейді. Позитивтік бақылау Көптеген бактериялық гендер позитивтік бақылаудың қарамағында болады. Гендер активтілігінің позитивтік бақылану механизмі нақты эффектор факторы бар кезде активтілікті күшейтіп РНҚ-полимеразаның промоторға жалғануы мен а-РНҚ-ның синтезделуін (транскрипцияны) жылдамдатады. Гендер активтілігін позитивті бақылауға мысал ретінде өзімізге белгілі E.coli-дің Lac –оперонын алайық. Позитивтік бақылаудың мәні мынада, жасушада глюкоза мен галактоза бар кезде Lac-оперон активті емес, сондықтан Lac – а-РНҚ-ның синтезі жүрмейді, β- галактозидаза ферменті де синтезделмейді. зделмейді
lac-оперонның позитивті реттелуі
Лактозалық і-репрессорын активсіздендіру үшін ортада тек глюкозаның болғаны жеткіліксіз. Глюкозаға қосымша элемент циклдық АМФ-катаболизмдік активтендіруші белок (КАБ)-тың болуы қажет. Олай болса жасушада Lac – а-РНҚ-ның транскрипциясы ол ортада индуктор ретінде глюкоза және циклдық (ц)АМФ-КАБ бар кезде жүреді. Циклдық (ц) АМФ-КАБ –тың активтілігі глюкозаның концентрациясына байланысты. Глюкоза Lac – оперонның активтілігіне жанама ингибиторлық (тоқтататындай) әсер етеді. Циклдық АМФ-тың кішігірім молекулалары ішек таяқшасынан басқа көптеген бактерияларда да кездеседі, олардың синтезделуі аденил – циклаза ферментінің көмегімен жүреді, оның жасушадағы концентрациясы жанама түрде глюкозаның метаболизмімен реттеледі. Егерде бактерия өсетін ортада глюкоза жеткілікті болатын болса онда цАМФ концентрациясы төмен болады, ал ортада глюкозаның метаболизміне қатыспайтын глицерол немесе көмірсулардың басқада көзідері болатын болса цАМФ концентрациясы жоғарылайды. цАМФ концентрациясының бақылану механизмі толық ашылған жоқ. Lac – оперонының активтілігінің реттелуінде цАМФ маңызды. Сонымен қатар ішек таяқшасының жасушасында Lac – оперонының активтілігін реттеуге қатынасатын катаболизмдік активтендіруші белок (КАБ) «сrp» генінің өнімі болып табылады. Осы сrp немесе аденил – циклаза гендерінің мутацияға ұшырауы Lac – а-РНҚ-ны синтездеу қабілетінен айырады, сондықтан Lac а-РНҚ синтезінің басталуы үшін жасушада осы екі геннің (цАМФ және КАБ) өнімдерінің болуы қажет. КАБ және ц АМФ бірігіп бірге Lac- жүйесінің активті реттеуші элементі болып табылады. Олар транскрипцияның басталуы үшін ДНК тізбегінің промоторлар қатарымен байланысуы қажет.Олай болса цАМФ және КАБ кешені репрессорға (негативті реттелуге) қарама-қарсы қасиет көрсететін позитивтік реттеуші болып табылады. Бұл Lac- оперонының активтілігі бір-біріне тәуелсіз позитивті және негативті механизмдермен реттелетіндігін көрсетеді.
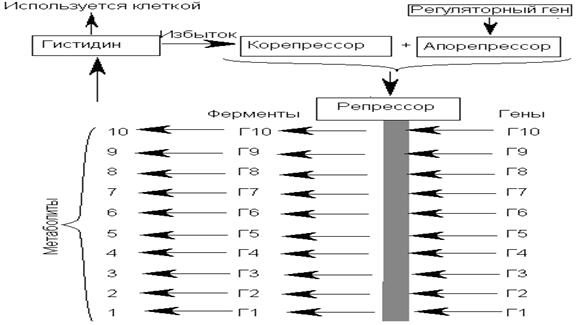
Репрессибельдік жүйелер Жасушадағы кейбір ферменттердің мөлшері олар синтездейтін соңғы өнімдердің концентрациясына байланысты өзгеріп отырады. Егер жасушада соңғы өнімдердің мөлшері жоғарылап, артып кетсе ол ферменттердің синтезі тоқталады. Ондай ферменттерді репрессибельдік ферменттер деп, ал соңғы өнімдерін – корепрессор деп атайды. Реттеуші ген апорепрессор деп аталатын затты синтездейді. Апорепрессор корепрессормен қосылып активті қызмет атқаратын репрессор молекуласын түзейді. Репрессор синтездік үдерістерге қатынасатын барлық гендердің активтілігін тежейді. Мысалы, Salmonella typhimurium бактериясында гистидин аминқышқылының концентрациясы жоғары ортада гистидин корепрессор ретінде әсер етеді. Ол корепрессор ретінде гистидиннің синтезіне қатынасатын барлық 10 ферменттің түзілуін тоқтатады. Мұндай репрессия типін үйлесімдік (координациялық) репрессия деп атайды. Репрессияның бұл типін координациялау репрессиясы деп атайды. Salmonellatyphimurium-ның координацияланған репрессиясы деп атайды. Трансляциялық бақылау Прокариоттық гендер активтілігінің трансляциялық деңгейде реттелуі а-РНҚ-ның өмір сүру ұзақтығына тікелей байланысты. А-РНҚ-ның ферменттік ыдырауы оның 51-ұшынан басталып 31-ұшында аяқталады, яғни а-РНҚ-ның бірінші болып синтезделген ұшы, бірінші болып ыдырайды. Ішек таяқшасының а-РНҚ-сының орташа өмір сүру ұзақтығы -2 минутке жуық. А-РНҚ-ның ферменттік ыдырау жылдамдығына оның 51-ұшында орналасқан арнайы нуклеотидтар жүйесі де әсер етуі мүмкін. А-РНҚ-ның өмір сүру ұзақтығы сонымен қатар оның трансляциясына қатынасатын бос рибосомалардың санына байланысты ауытқиды. Осы себепті бактерияларда түрлі белоктар синтезі жылдамдығының түрліше болуы трансляция жылдамдығымен емес сол жасушадағы рибосомалар санымен анықталады. Мысалы, ішек таяқшасының (E. coli) лактозалық жүйесіндегі жалпы бір оператормен бақыланатын құрылымдық гендердегі үш цистрон үш ферментті синтездейтіні айтылды. Олар: а) β-галактозидаза; ә) галактозид-пермеаза; б) галактозид-ацетилаза. Бұл белоктардың синтезделу арақатынасы 1:1/2:1/5-ке тең. Бұл арақатынас осы үш белоктың трансляциясы жүретін полицистрондық а-РНҚ-ның 51-ұшынан олардың орналасу арақашықтығын бейнелейді. Гендер неғұрлым полицистронды а-РНҚ - 51-ұшынан неғұрлым алыс орналасқан сайын синтезделетін белоктың мөлшері де азая түседі. Рибосомалардың а-РНҚ-ғы түрлі старттық нүктелерге жалғануы оперондағы құрылымдық Z, Y, A гендері синтездейтін белоктардың сан мөлшерінің түрліше болуына әсерін тигізеді деген жорамал айтылады. Посттрансляциялық бақылау Гендер активтілігі белоктардың синтезі аяқталғаннан кейін де реттелуі мүмкін. Бақылаудың бұл түрін гендер активтілігінің посттрансляциялық бақылануы деп атайды. Гендер активтілігінің кері байланысу ұстанымы бойынша басылуы ферменттің синтезіне емес, оның активтілігіне әсер ететін реттеуші механизм болып табылады. Мұнда биосинтез нәтижесінде түзілген концентрациясы жоғары соңғы өнім бірінші синтезделген ферментпен байланысып метаболизмдік үдерістің жүруіне жағдай туғызады. Соңғы өнім + бірінші фермент кешені ферменттің төртінші реттік құрылымын өзгертіп, оның катализдік сайтын активсіздендіреді. Фермент құрылымының өзгеруі оның өзінің активтілігін тоқтатады және соңғы өнім мен аралық ыдырау заттарының артық мөлшерде синтезделуіне кедергі жасайды. Мысалы, ішек таяқшасында изолейциннің синтезделу үдерісін зерттеу барысында (Umbеrger, 1961) изолейцинді (треонин алмасудағы соңғы өнім) жасанды ортаға енгізгенде, сол мезетте треониннің изолейцинге айналу үдерісі тоқталған. Изолейциннің мөлшері шамадан тыс артқан жағдайда жасушада бұл аминқышқылының синтезі тоқталып, экзогендік изолейцинді пайдалану басымырақ жүрген. Треониннің изолейцинге айналу процесіне қатынасатын 5 фермент артық мөлшердегі изолейцинмен өзара әрекеттесуі барысында олардың әрқайсысының синтезі де тоқталады.
6.5. Оқыту әдістері:студенттердің сабақтың мәнін, мақсатын және міндеттерін, түсінгенін қысқа, анық, ойын жүйелі жеткізу және түсінген материалын сызба нұсқа, кесте, сурет түрінде көрсету; тест қателіктерімен топпен жұмыс: есептер шығаруды көрсету және түсіндіру, сызбалар толтыруға бағыттай отырып ауызша сұрау.
6.6. Әдебиеттер: Негізгі: 6.6. 1. Муминов Т.А.,Куандыков Е.У. Молекулалық биология негіздері (дәрістер жинағы). Алматы: Эверо, 2009. С. 83-90. 6.6.2. Мушкамбаров Н.Н., Кузнецов С.Л. Молекулярная биология. М., 2003. С. 17-27. 6.6.3. Медициналықя биология және генетика. Оқу құралы под ред. проф. Куандыкова Е.У. Алматы, 2004. С.29-31. Қосымша: 6.6.4. Гинтер Е.К. Медицинская генетика. М.: Медицина, 2003. С. 28-50. 6.6. 5. Жимулев И.Ф. Общая и молекулярная генетика. Новосибирск: Сиб.унив. изд-во, 2007. С. 107-196. 6.6.6. Медицинская генетика: учеб. пособие/ Роберт Л.Ньюссбаум, Родерик Р, Мак-Иннес, Хантингтон Ф. Виллард: пер. с англ. А. Ш. Латыпова; под ред. акад. РАМН Н. П. Бочкова.- М.: ГЭОТАР-Медиа, 2010. 6.7. Бақылау: 6.7.1. Құзыреттілікті бағалау – білім. 6.7.1.2. Тақырыптың сұрақтары бойынша ауызша сұрау. 6.7.1.3. Тесттік бақылау – 10 сұрақтан 3 нұсқа. 6.7.1.4. Тақырып бойынша кәсіби терминдерді ағылшын және екінші тілде игеруін тексеру
6.7.2. Тәжірибелік дағды құзыреттілігін бағалау: 6.7.2.1. Сызбаларды салу: - оперонның моделі (Қуандықов Е.Ө., Аманжолова Л.Е. Молекулалық биология негіздері.б. 85, сурет 43, бірінші қатары); 6.7.2. Тәжірибелік дағды құзыреттілігін бағалау.Сызбаларды сызу: 6.7.2.1. - индуцебелді оперонның моделін; 6.7.2.2. – репрессибелді оперон 6.7.3.2. Есептерді шығару. РНҚ полимеразаны танитын сайты- ТАТААТ, -35 жүйелік ТТ ГАЦ А, репрессор – ақуызының оперондағы танитын сайты (оператор)-АТТТТАТ, құрылымдық гендер ТАЦ кодондарымен басталып АТЦ кодондарымен аяқталады. Гендер аралығында спейсерлік ТТТТ бөліктер орналасқан. а) осы ақпаратты пайдалана отырып, 3 құрылымдық геннен тұратын Lac–оперонының моделін құрастырыңыз: құрылымдық ген Z- 5 кодоннан тұрады, құрылымдық гендер Y және A - 4 кодоннан тұрады. б) транскрипция жүргізіңіз, қанша а-РНҚ түзіледі? в) трансляция жүргізіңіз, ДНҚ-ң осы бөлігінде қанша ақуыз кодталған? Олардың құрамын анықтаңыз.
6.7.2.3. Кесте толтыру:
Прокариот гендерінің реттелу деңгейлері
6.7.3. Бақылау сұрақтары: 6.7.3.1. Гендер белсенділігінің критериі болып не табылады? 6.7.3.2. Оперон деген не, прокариоттар гендерінің реттелуіндегі оның ролі неде? 6.7.3.3. Прокариоттар гендерінің белсенділігінің реттелу механизмдерінің бұзылуының медициналық зардаптары қандай? Үш тілдегі түсіндірме сөздер:
Құрастырушы: оқыт. Жумагул М.Ж.
№26 6.1. Тақырыбы: Эукариот гендерінің экспрессиясының реттелуі 6.2. Мақсаты: Студенттерде гендер экспрессиясының мәні, эукариот гендерінің белсенділігінің реттелуінің молекулалық механизмдері және гендер экспрессиясының бұзылуында патологиялық жағдайлардың дамуы мен пайда болу ролі жайлы білімін қалыптастыру. 6.3. Оқытудың міндеттері: - эукариоттар гендер белсенділігінің ерекшелігін оқыту - гендер транскрипциясы мен трансляциясының белсенділігінің ерекшелігінің ролін оқыту - студенттерде гендер белсенділігінің реттелуінің бұзылуының маңызы жайлы түсінікті қалыптастыру - ағзалар тіршілігінде фундаменталдық үдерісте әр түрлі гендер белсенділігінің ролін оқыту: онтогенез, морфогенез, гистогенез - студенттерде тірі ағза мен жасушаларда гендер белсенділігінің реттелуінің бұзылуына байланысты патологиялық өзгерістердің пайда болу маңызы жайлы түсінікті қалыптастыру 6.4. Тақырыптың негізгі сұрақтары: * 6.4.1. Гендерінің белсенділігінің реттелу түсінігі, мәні |
||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2018-05-10; просмотров: 1776. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |