|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
Ацетил-КоА: источники и основные пути использования в тканях. Компартментализация обмена ацетил-КоА.
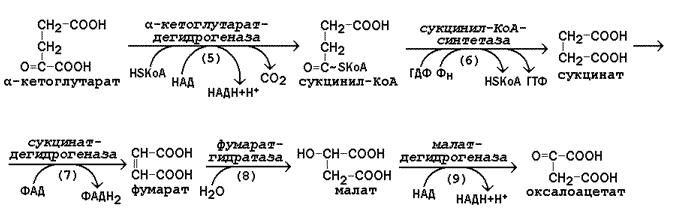
Дальнейшем пируват, а также жирные кислоты и некоторые аминокислоты окисляются до ацетильного остатка, связанного с коэнзимом А (ацетил-КоА). Эти реакции протекают уже в митохондриях клетки. Пируват и ацетил-КоА, находящиеся на пересечении нескольких метаболических путей, можно отнести кключевым или узловым метаболитам. 39. Цикл трикарбоновых кислот: последовательность реакций, характеристика ферментов. Амфиболическая функция цитратного цикла. Связь с обменом углеводов, жиров и белков. Реакции цикла Кребса относятся к третьей стадии катаболизма питательных веществ и происходят в митохондриях клетки. Эти реакции относятся к общему пути катаболизма и характерны для распада всех классов питательных веществ (белков, липидов и углеводов). Главной функцией цикла является окисление ацетильного остатка с образованием четырёх молекул восстановленных коферментов (трёх молекул НАДН и одной молекулы ФАДН2), а также образование молекулы ГТФ путём субстратного фосфорилирования. Атомы углерода ацетильного остатка выделяются в виде двух молекул СО2. 13.4.2.Цикл Кребса включает 8 последовательных стадий, обращая особое внимание на реакции дегидрирования субстратов:
Рисунок 13.6. Реакции цикла Кребса, включая образование α-кетоглутарата а) конденсация ацетил-КоА с оксалоацетатом, в результате которой образуется цитрат (рис.13.6, реакция 1); поэтому цикл Кребса называют также цитратным циклом. В этой реакции метильный углерод ацетильной группы взаимодействует с кетогруппой оксалоацетата; одновременно происходит расщепление тиоэфирной связи. В реакции освобождается КоА-SH, который может принять участие в окислительном декарбоксилировании следующей молекулы пирувата. Реакцию катализирует цитратсинтаза, это – регуляторный фермент, он ингибируется высокими концентрациями НАДН, сукцинил-КоА, цитрата.  б) превращение цитрата в изоцитрат через промежуточное образование цис-аконитата. Образующийся в первой реакции цикла цитрат содержит третичную гидроксильную группу и не способен окисляться в условиях клетки. Под действием фермента аконитазы идёт отщепление молекулы воды (дегидратация), а затем её присоединение (гидратация), но другим способом (рис.13.6, реакции 2-3). В результате данных превращений гидроксильная группа перемещается в положение, благоприятствующее её последующему окислению. в) дегидрирование изоцитрата с последующим выделением молекулы СО2 (декарбоксилированием) и образованием α-кетоглутарата (рис. 13.6, реакция 4). Это – первая окислительно-восстановительная реакция в цикле Кребса, в результате которой образуется НАДН. Изоцитратдегидрогеназа, катализирующая реакцию, - регуляторный фермент, активируется АДФ. Избыток НАДН ингибирует фермент.
Рисунок 13.7. Реакции цикла Кребса, начиная с α-кетоглутарата. г) окислительное декарбоксилирование α-кетоглутарата, катализируется мультиферментным комплексом (рис. 13.7, реакция 5), сопровождается выделением СО2 и образованием второй молекулы НАДН. Эта реакция аналогична пируватдегидрогеназной реакции. Ингибитором служит продукт реакции – сукцинил-КоА. д) субстратное фосфорилирование на уровне сукцинил-КоА, в ходе которого энергия, освобождающаяся при гидролизе тиоэфирной связи, запасается в форме молекулы ГТФ. В отличие от окислительного фосфорилирования, этот процесс протекает без образования электрохимического потенциала митохондриальной мембраны (рис. 13.7, реакция 6). е) дегидрирование сукцината с образованием фумарата и молекулы ФАДН2 (рис. 13.7, реакция 7). Фермент сукцинатдегидрогеназа прочно связан с внутренней мембраной митохондрии. ж) гидратация фумарата, в результате чего в молекуле продукта реакции появляется легко окисляемая гидроксильная группа (рис. 13.7, реакция 8). з) дегидрирование малата, приводящее к образованию оксалоацетата и третьей молекулы НАДН (рис.13.7, реакция 9). Образующийся в реакции оксалоацетат может вновь использоваться в реакции конденсации с очередной молекулой ацетил-КоА (рис. 13.6, реакция 1). Поэтому данный процесс носит циклический характер. 13.4.3. Таким образом, в результате описанных реакций подвергается полному окислению ацетильный остаток СН3-СО-. Количество молекул ацетил-КоА, превращаемых в митохондриях в единицу времени, зависит от концентрации оксалоацетата. Основные пути увеличения концентрации оксалоацетата в митохондриях (соответствующие реакции будут рассмотрены позднее): а) карбоксилирование пирувата – присоединение к пирувату молекулы СО2 с затратой энергии АТФ; б) дезаминирование или трансаминирование аспартата – отщепление аминогруппы с образованием на её месте кетогруппы. 13.4.4. Некоторые метаболиты цикла Кребса могут использоваться для синтеза структурных блоков для построения сложных молекул. Так, оксалоацетат может превращаться в аминокислоту аспартат, а α–кетоглутарат – в аминокислоту глутамат. Сукцинил-КоА принимает участие в синтезе гема – простетической группы гемоглобина. Таким образом, реакции цикла Кребса могут участвовать как в процессах катаболизма, так и анаболизма, то есть цикл Кребса выполняет амфиболическую функцию (см. 13.1). 40. Окисление НАДН2 и ФАДН2 в митохондриях. Характеристика основных компонентов дыхательной цепи. Ферментные комплексы. Дегидрирование субстратов и окисление водорода как источник энергии для синтеза АТФ. Роль АТФ в организме. В пируватдегидрогеназной реакции и в цикле Кребса происходит дегидрирование (окисление) субстратов (пируват, изоцитрат, α-кетоглутарат, сукцинат, малат). В результате этих реакций образуются НАДН и ФАДН2. Эти восстановленные формы коферментов окисляются в митохондриальной дыхательной цепи. Окисление НАДН и ФАДН2, протекающее сопряжённо с синтезом АТФ из АДФ и Н3РО4 называется окислительным фосфорилированием. Схема строения митохондрии показана на рисунке 14.1. Митохондрии представляют собой внутриклеточные органеллы, имеющие две мембраны: наружную (1) и внутреннюю (2). Внутренняя митохондриальная мембрана образует многочисленные складки – кристы (3). Пространство, ограниченное внутренней митохондриальной мембраной, носит название матрикс (4), пространство, ограниченное наружной и внутренней мембранами, - межмембранное пространство Дыхательная цепь– последовательная цепь ферментов, осуществляющая перенос ионов водорода и электронов от окисляемых субстратов к молекулярному кислороду – конечному акцептору водорода. В ходе этих реакций выделение энергии происходит постепенно, небольшими порциями, и она может быть аккумулирована в форме АТФ. Локализация ферментов дыхательной цепи – внутренняя митохондриальная мембрана. Дыхательная цепь включает четыре мультиферментных комплекса I. НАДН-KoQ-редуктаза (содержит промежуточные акцепторы водорода: флавинмононуклеотид и железосерные белки). II. Сукцинат-KoQ-редуктаза (содержит промежуточные акцепторы водорода: ФАД и железосерные белки). III. KoQН2-цитохром с-редуктаза (содержит акцепторы электронов: цитохромы b и с1, железосерные белки). IV. Цитохром с-оксидаза (содержит акцепторы электронов: цитохромы а и а3, ионы меди Cu2+). 14.1.3. В качестве промежуточных переносчиков электронов выступают убихинон (коэнзим Q) и цитохром с. Убихинон (KoQ) – жирорастворимое витаминоподобное вещество, способен легко диффундировать в гидрофобной фазе внутренней мембраны митохондрий. Биологическая роль коэнзима Q – перенос электронов в дыхательной цепи от флавопротеинов (комплексы I и II) к цитохромам (комплекс III). Цитохром с – сложный белок, хромопротеин, простетическая группа которого – гем – содержит железо с переменной валентностью (Fe3+ в окисленной форме и Fe2+ в восстановленной форме). Цитохром сявляется водорастворимым соединением и располагается на периферии внутренней митохондриальной мембраны в гидрофильной фазе. Биологическая роль цитохрома с – перенос электронов в дыхательной цепи от комплекса III к комплексу IV. 14.1.4. Промежуточные переносчики электронов в дыхательной цепи расположены в соответствии с их окислительно-восстановительными потенциалами. В этой последовательности способность отдавать электроны (окисляться) убывает, а способность присоединять электроны (восстанавливаться) возрастает. Наибольшей способности отдавать электроны обладает НАДН, наибольшей способностью присоединять электроны – молекулярный кислород. Механизм синтеза АТФ описывает хемиосмотическая теория (автор - П. Митчелл). Согласно этой теории, компоненты дыхательной цепи, расположенные во внутренней митохондриальной мембране, в ходе переноса электронов могут «захватывать» протоны из матрикса митохондрий и передавать их в межмембранное пространство. При этом наружная поверхность внутренней мембраны приобретает положительный заряд, а внутренняя – отрицательный, т.е. создаётся градиент концентрации протонов с более кислым значением рН снаружи. Так возникает трансмембранный потенциал (ΔµН+). Существует три участка дыхательной цепи, на которых он образуется. Эти участки соответствуют I, III и IV комплексам цепи переноса электронов. Энергия, аккумулированная в форме АТФ, используется в организме для обеспечения разнообразных биохимических и физиологических процессов. Запомните основные примеры использования энергии АТФ: 1) синтез сложных химических веществ из более простых (реакции анаболизма); 2) сокращение мышц (механическая работа); 3) образование трансмембранных биопотенциалов; 4) активный транспорт веществ через биологические мембраны.
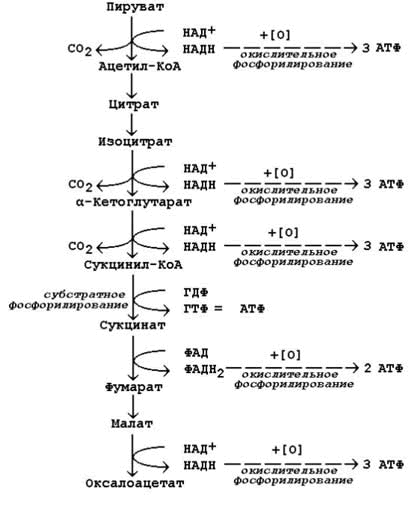
Рисунок 10.5. Строение неконъюгированных (а) и конъюгированных (б) желчных кислот. 15.1.4.Желчные кислоты обладают амфифильными свойствами: гидроксильные группы и боковая цепь гидрофильны, циклическая структура гидрофобна. Эти свойства обусловливают участие желчных кислот в переваривании липидов: 1) желчные кислоты способны эмульгировать жиры, их молекулы своей неполярной частью адсорбируются на поверхности жировых капель, в то же время гидрофильные группы вступают во взаимодействие с окружающей водной средой. В результате снижается поверхностное натяжение на границе раздела липидной и водной фаз, вследствие чего крупные жировые капли разбиваются на более мелкие; 2) желчные кислоты наряду с колипазой желчи участвуют в активировании панкреатической липазы, сдвигая её оптимум рН в кислую сторону; 3) желчные кислоты образуют с гидрофобными продуктами переваривания жиров водорастворимые комплексы, что способствует их всасыванию в стенку тонкого кишечника. Желчные кислоты, проникающие в процессе всасывания вместе с продуктами гидролиза в энтероциты, через портальную систему поступают в печень. Эти кислоты могут повторно секретироваться с желчью в кишечник и участвовать в процессах переваривания и всасывания. Такая энтеро-гепатическая циркуляция желчных кислот может осуществляться до 10 и более раз в сутки. 41. Сопряжение окисления с фосфорилированием в дыхательной цепи. Н+-АТФсинтетаза мембран митохондрий. Коэффициент Р/О. Разобщение дыхания и фосфорилирования. Гипоэнергетические состояния. Степень сопряжённости окисления и фосфорилирования в митохондриях характеризует коэффициент фосфорилирования (Р/О). Он равен отношению количества молекул неорганического фосфата (Н3РО4), перешедшего в АТФ, к количеству атомов потреблённого кислорода (О2). Например, если донором водорода для дыхательной цепи является молекула НАДН, то электроны от донора (НАДН) к акцептору (кислород) проходят 3 участка сопряжения окисления и фосфорилирования (I, III и IV ферментные комплексы дыхательной цепи). Таким образом, максимально может образоваться 3 молекулы АТФ (3 АДФ + 3 Н3РО4 → 3 АТФ). Затрачивается 1 атом кислорода (2 Н + О → Н2О). Значение коэффициента Р/О = 3/1 = 3. Если донором водорода будет молекула ФАДН2, то электроны в дыхательной цепи проходят 2 участка сопряжения окисления и фосфорилирования (III и IV ферментные комплексы дыхательной цепи). Таким образом, максимально может образоваться 2 молекулы АТФ (2 АДФ + 2 Н3РО4 → 2 АТФ). Затрачивается, как и в предыдущем случае, 1 атом кислорода (2 Н + О → Н2О). Значение коэффициента Р/О = 2/1 = 2. 14.2.2.Более сложный пример расчёта коэффициента фосфорилирования – при окислении пирувата до конечных продуктов - показан на рисунке 14.5. В этом метаболическом пути происходит дегидрирование 4 субстратов (пирувата, изоцитрата, α-кетоглутарата и малата) с образованием НАДН и одного субстрата (сукцината) с образованием ФАДН2. Восстановленные коферменты окисляются в дыхательной цепи, и в сопряжённых реакциях фосфорилирования образуется (4×3 АТФ + 1×2 АТФ)=14 молекул АТФ. Ещё 1 молекула АТФ (ГТФ) образуется в реакции субстратного фосфорилирования на уровне сукцинил-КоА. Таким образом, при полном окислении 1 молекулы пирувата образуется 15 молекул АТФ (из них 14 - путём окислительного фосфорилирования). Чтобы рассчитать количество потреблённого кислорода, нужно знать число реакций дегидрирования на данном участке метаболического пути. Для окисления каждой восстановленной формы кофермента необходим 1 атом кислорода (см. выше). Следовательно, в нашем примере потребляется 5 атомов кислорода. Значение коэффициента Р/О будет равно 14/5 = 2,8. Перенос электронов в дыхательной цепи не во всех случаях протекает сопряжённо с фосфорилированием АДФ. Состояние, при котором окисление субстратов в дыхательной цепи происходит, но АТФ при этом не образуется, называется свободным (нефосфорилирующим) окислением. Энергия, выделяемая при окислении, рассеивается в виде теплоты. В физиологических условиях свободное окисление может служить одним из механизмов терморегуляции. В организме человека и некоторых животных имеется особая ткань – бурый жир, содержащий митохондрии, приспособленные для выработки теплоты. Много бурого жира у новорождённых, в последующие периоды жизни его количество уменьшается. В митохондриях бурого жира содержание дыхательных ферментов значительно выше, чем ферментов, осуществляющих фосфорилирование АДФ, поэтому в них преобладают процессы свободного окисления. Разобщение процессов окисления и фосфорилирования в митохондриях может иметь место при некоторых патологических состояниях. Основными симптомами таких состояний могут быть быстрая утомляемость, повышенная температура тела, снижение массы тела, несмотря на повышенный аппетит, учащение дыхания и сердцебиения.
Рисунок 14.5.Расчёт энергетического баланса реакций окислительного декарбоксилирования пирувата и цикла Кребса. 14.3.2.Разобщение процессов окисления и фосфорилирования может быть вызвано действием ряда веществ, как природных, так и синтетических. Механизм действия этих веществ заключается в том, что они являются переносчиками протонов через мембрану. Вещества, разобщающие окисление и фосфорилирование, можно разделить на протонофоры и ионофоры. Протонофоры представляют собой слабые гидрофобные органические кислоты, которые в форме аниона (R-COO-) связывают протоны в межмембранном пространстве, диффундируют через мембрану и диссоциируют в матриксе с образованием протонов. К этой группе относятся, например, свободные жирные кислоты, гормоны щитовидной железы, салицилаты, дикумарол, 2,4-динитрофенол (см. рисунок 14.6).
Рисунок 14.6.Механизм действия 2,4-динитрофенола. |
||
|
|
Последнее изменение этой страницы: 2018-05-10; просмотров: 535. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |