|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
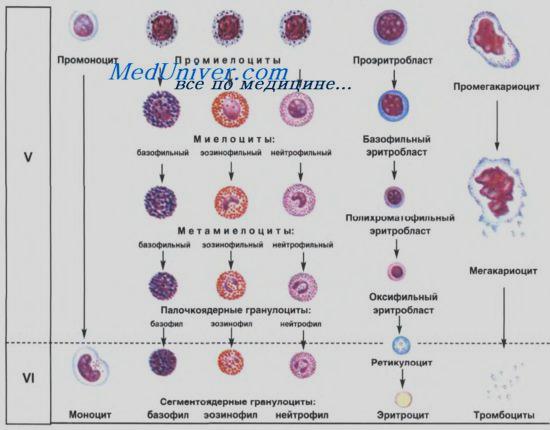
Клеточные основы кооперации в иммунных реакциях. Взаимодействие клеток при иммунном ответе.Стр 1 из 3Следующая ⇒ Гранулоцитопоэз. Образование гранулоцитов происходит в миелоидной ткани красного костного мозга. Исходная стволовая клетка превращается в мультипотентную клетку — предшественник миелопоэза (КОЕ-ГЭММ) и далее под воздействием колониестимулирующих факторов дифференцируется в общую родоначальную клетку для гранулоцитов и моноцитов (КОЕ-ГМн). В дальнейшем в результате дивергенции возникают родоначальные клетки для гранулоцитов (КОЕ-Гн), которые дифференцируются в идентифицируемые миелобласты (IV-й класс клеток). В ряду дальнейшей клеточной дифференцировки (V-й класс клеток) различают стадии: промиелоцита, миелоцита, метамиелоцита. Начиная со стадии промиелоцита, клетки подразделяются на 3 разновидности: нейтрофильные, эозинофильные, базофильные. Более отчетливо это подразделение можно провести на стадии миелоцитов, когда в клетках накапливается достаточное количество специфической зернистости. До стадии миелоцитов включительно клетки гранулоцитопоэза делятся митозом. Метамиелоциты митозом уже не делятся. В этих клетках ядро приобретает вначале палочковидную, а затем сегментированную форму. Общее направление дифференцировки клеток гранулопоэза характеризуется: постепенным уменьшением размеров клетки, снижением базофилии цитоплазмы, появлением в цитоплазме специфических гранул, уменьшением размеров ядра, появлением сегментированности ядра и его уплотнением, сдвигом ядерно-цитоплазменного отношения в сторону преобладания размеров цитоплазмы над размерами ядра.
 9 Клеточные основы кооперации в иммунных реакциях. Взаимодействие клеток при иммунном ответе. Реактивное свойство организма, которое проявляется на клеточном уровне организации живого и заключается в распозновании "своих и чужих" клеток, обеспечивается Т-лимфоцитами. Молекулярные механизмы "узнавания" заключаются в том, что практически все клетки организма позвоночных экспрессируют молекулу гистосовместимости, которая находится на поверхности клетки и доступна для опознавания специализированными клетками иммунной системы. В связи с тем, что молекула в составе клетки одного организма, будучи внесенная в другой, вызывает реакцию отторжения, она получила название антиген гистосовместимости, или главная молекула гистосовместимости. Последняя бывает двух классов — I и II. Молекула гистосовместимости 1-го класса является гликопротеином. Она состоит из трех частей: а) трансмембранного белка (тяжелой цепи), который пронизывает плазмолемму клетки и имеет внутриклеточную, цитоплазматическую и внеклеточную части (домены); б) белка малой молекулярной массы (эпитопа), расположенного между петлями внеклеточной части трансмембранного белка; в) периферического белка. Гены, кодирующие молекулярную организацию трансмембранного белка, расположены на 6-й хромосоме в области, которая получила название главный комплекс гистосовместимости (англ. — МНС — Major Histocompatibility Complex).
Цитотоксические лимфоциты имеют рецепторы, с помощью которых осуществляется мониторинг практически всех клеток организма (за исключеним тех, которые отделены специальными гематотканевыми гистионами). Они имеют специфические рецепторы — Т-клеточные, с помощью которых взаимодействуют с эпитопом тканевой клетки, и особые рецепторы, представленные поверхностными гликопротеинами, которые выявляются моноклональными антителами CD8 (от англ. cluster of differentiation по номенклатуре, принятой в иммуноцитохимии рецепторов). Если Т-лимфоциты маркируются моноклональным антителом CD8, то они называются CD8+ T-лимфоцитами, цитотоксическими, или киллерами. Последние связываются только с молекулой гистосовместимости 1-го класса. В случае обнаружения чужеродного эпитопа происходит молекулярное взаимодействие в области контакта рецепторов, что является сигналом к запуску механизмов разрушения клетки, синтезирующей чужеродный белок. Это проявляется в секреции в межклеточное пространство белков — перфоринов, которые в виде трансмембранных белков встраиваются в плазмолемму чужеродной клетки, формируют поры, приводящие к нарушению гомеостаза, набуханию и распаду клетки. Кроме того, цитотоксические лимфоциты способны активизировать своими биологически активными веществами ферменты апоптоза как при проникновении их в клетку через поры, так и через рецепторы чужеродной клетки к индукторам апоптоза. Существуют и другие механизмы цитотоксического действия данной субпопуляции лимфоцитов на мишени. В отличие от реакций клеточного типа существует другой тип реакции иммунных клеток, выражающийся в выработке антител в жидкие среды организма. Рецепторная система клеток (макрофагов, В- и Т-лимфоцитов), участвующих в иммунных реакциях данного типа, представлена молекулами гистосовместимости II-го класса. Последняя состоит из двух трансмембранных белков, между петлями которых располагается фрагмент антигена — эпитоп, или антигенная детерминанта. В отличие от описанного выше процесса возникновения эпитопа в составе молекулы гистосовместимости 1-го класса, во втором случае эпитоп появляется как результат двух процессов: а) фагоцитоза антигена макрофагом (антигенпредставляющей клеткой), внутриклеточного взаимодействия антигена с лизосомами макрофага и возврата фрагмента антигена в виде эпитопа на клеточную поверхность; б) рецепторно-опосредованного эндоцитоза В-лимфоцитами растворенного антигена, его модификации и возврата в виде эпитопа на клеточную поверхность. Следовательно, принципиальные отличия в организации молекулы гистосовместимости II-го класса заключаются в том, что не все клетки синтезируют данный класс молекул, а эпитоп имеет внеклеточное происхождение. С молекулой II-го класса могут взаимодействовать только те лимфоциты, которые имеют на своей поверхности другой — специальный рецептор класса CD4. К ним относятся СD+4Т-лимфоциты, или Т-хелперы. Последние вместе с антигенп-редставляющими клетками и В-лимфоцитами отвечают за формирования иммунной реакции, которая проявляется в синтезе иммуноглобулинов, или антител, циркулирующих в крови и тканевой жидкости (гуморальный иммунитет). Межклеточные взаимодействия протекают следующим образом. Т-хелпер с помощью двух своих рецепторов — Т-клеточного (молекулы II-го класса) и CD4 связывается с молекулой II-го класса гистосовместимости макрофага (антигенпредставляющей клетки). Если Т-клеточный рецептор с одной стороны и эпитоп молекулы II-го класса макрофага с другой стороны не комплементарны, то происходит активация и пролиферации Т-хелпера. Аналогично происходит взаимодействие хелпера с В-лимфоцитом. Если эпитоп молекулы II-го класса В-лимфоцита не соответствует Т-клеточному рецептору хелпера, то последний вырабатывает лимфокины, которые вызывают пролиферацию и дифференцировку В-лимфоцита в плазматические клетки. Последние специализированы на синтезе в большом количестве клеточных рецепторов к конкретному антигену и секреции рецепторов в виде иммуноглобулинов в кровь, лимфу, тканевую жидкость. В свою очередь рецепторы связывают циркулирующие в жидкостях организма антигены, предупреждают дальнейшее развитие заболевания. Итак, иммуноглобулины — это секретируемые плазматическими клетками клеточные рецепторы к определенным антигенам. СD4+Т-лимфоциты называются помощниками (хелперами), так как без их участия невозможна активация, пролиферация и дифференцировка В-лимфоцитов в плазматические клетки. Таким образом, иммунный комплекс органов действует как целое. Благодаря взаимодействию с эндокринной, нервной системами и местными факторами регуляции, происходит пролиферация и дифференцировка лимфоидных клеток, упорядоченная их миграция, осуществляемая через кровь и лимфу. Медленная миграция клеток характерна для стволовых клеток, Т- и В-лимфоцитов, заселяющих периферические органы иммуногенеза. Быстрая миграция характерна для рециркулирующих долгоживущих лимфоцитов памяти. Количество рециркулирующих клеток составляет 1010. Они мигрируют из крови в периферические органы иммуногенеза и обратно. Рециркуляция Т-лимфоцитов обеспечивает поддержание иммунного гомеостаза. Лимфоцитопоэз и иммуноцитопоэз. Лимфоидная ткань у человека имеется в составе лимфатических узлов, селезенки, миндалин, аппендикса и в других лимфоидных образованиях по ходу пищеварительного тракта. В лимфоидной ткани происходит лимфопоэз. Исходными клетками лимфопоэза являются стволовые клетки красного костного мозга. Через стадию мультипотентных клеток (КОЕ-Л) они дифференцируются в родоначальные про-Т- и про-В-лимфобласты и далее в Т- и В-лимфобласты, Т- и В-пролимфоциты и Т- и В-лимфоциты. В лимфоцитопоэзе в тимусе возникают субпопуляции Т-клеток с различными рецепторами (так называемая антигеннезависимая пролиферация и дифференцировка). Т-лимфоциты участвуют в формировании клеточного иммунитета. Другой ряд дифференцировки в лимфопоэзе приводит к образованию из В-лимфоцитов через стадии плазмобласта и проплазмоцита — плазматических клеток (плазмоцитов). Эти клетки вырабатывают антитела, обеспечивая гуморальный иммунитет. Подробнее образование иммунокомпетентных клеток и их участие в развитии воспаления рассматриваются ниже. Из лимфобластов образуются большие, средние и малые лимфоциты. Этот ряд дифференцировки сопровождается уменьшением размеров клеток, уплотнением ядер, снижением митотической активности. Малые лимфоциты способны к "бласттрансформации" — своеобразной дедифференцировке с последующей повторной их дифференцировкой. Явление бласттрансформации открыто А.А. Максимовым (1902).
Костные ткани — полидифферонные ткани и состоят из клеток различной гистогенетической детерминации (остеобластов, остеоцитов и остеокластов) и очень плотного межклеточного вещества, содержащего большое количество минеральных солей. Костные ткани выполняют опорную функцию. Они входят в качестве главного структурного компонента в состав скелета. Благодаря высокому содержанию минеральных солей (до 65-70% сухой массы) костные ткани активно участвуют в регуляции минерального обмена. Между костными и кроветворными тканями складываются особые взаимодействия, обеспечивающие благоприятное микроокружение для пролиферации и дифференцировки клеток крови. По степени упорядоченности расположения коллагеновых волокон, которые в костной ткани называются оссеиновыми, различают ретикулофиброзную (грубоволокнистую) и пластинчатую костные ткани. Кроме того, существует дентиноидная костная ткань (дентин зуба), а также цемент зуба. Остеобласты (от греч. osteon — кость, blastos — зачаток), — это молодые клетки, создающие костную ткань. В кости они встречаются только в надкостнице. Они способны к пролиферации. В образующейся кости остеобласты покрывают почти непрерывным слоем всю поверхность развивающейся костной балки. Форма остеобластов бывает различной: кубической, пирамидальной или угловатой. Размер их тела около 15—20 мкм. Ядро округлой или овальной формы, часто располагается эксцентрично, содержит одно или несколько ядрышек. В цитоплазме остеобластов хорошо развиты гранулярная эндоплазматическая сеть, митохондрии и аппарат Гольджи. В ней выявляются в значительных количествах РНК и высокая активность щелочной фосфатазы. Остеоциты (от греч. osteon — кость, cytus — клетка) — это преобладающие по количеству зрелые (дефинитивные) клетки костной ткани, утратившие способность к делению. Они имеют отростчатую форму , компактное, относительно крупное ядро и слабобазофильную цитоплазму. Органеллы развиты слабо. Наличие центриолей в остеоцитах не установлено. Костные клетки лежат в костных лакунах, которые повторяют контуры остеоцита. Длина полостей колеблется от 22 до 55 мкм, ширина — от 6 до 14 мкм. Канальцы костных лакун заполнены тканевой жидкостью, анастомозируют между собой и с периваскулярными пространствами сосудов, заходящих внутрь кости. Обмен веществ между остеоцитами и кровью осуществляется через тканевую жидкость этих канальцев. Остеокласты (от греч. osteon — кость и clastos — раздробленный), - это клетки гематогенной природы, способные разрушать обызвествленный хрящ и кость. Диаметр их достигает 90 мкм и более, и они содержат от 3 до нескольких десятков ядер. Цитоплазма слабобазофильна, иногда оксифильна. Остеокласты располагаются обычно на поверхности костных перекладин. Та сторона остеокласта, которая прилежит к разрушаемой поверхности, богата цитоплазматическими выростами (гофрированная каемка); она является областью синтеза и секреции гидролитических ферментов. По периферии остеокласта находится зона плотного прилегания клетки к костной поверхности, которая как бы герметизирует область действия ферментов. Эта зона цитоплазмы светлая, содержит мало органелл, за исключением микрофиламентов, состоящих из актина. Периферический слой цитоплазмы над гофрированным краем содержит многочисленные мелкие пузырьки и более крупные — вакуоли. Полагают, что остеокласты выделяют СО2 в окружающую среду, а фермент карбоангидраза способствует образованию угольной кислоты (Н2СО3) и растворению кальциевых соединений. Остеокласт богат митохондриями и лизосомами, ферменты которых (коллагеназа и другие протеазы) расщепляют коллаген и протеогликаны матрикса костной ткани. Считается, что один остеокласт может разрушить столько кости, сколько создают 100 остеобластов за это же время. Функции остеобластов и остеокластов взаимосвязаны и регулируются гормонами, простагландинами, функциональной нагрузкой, витаминами и др.
11
Актиновые миофиламенты, содержащие белок актин, образуют изотропные диски (I). Это светлые, не обладающие двойным лучепреломлением диски. В центре дисковIпроходитZ-линия — телофрагма.Эта линия делит дискI на два полудиска. В области Z-линий расположены так называемые триады. Триады состоят из трубчатых элементов — Т-трубочек, образованных вдавлением плазмолеммы внутрь мышечного волокна. По этим трубочкам нервный импульс поступает к миофибриллам. В каждой триаде одна Т- Выделяют несколько последовательных этапов запуска и осуществления мышечного сокращения. 5. Потенциал действия проводится вдоль мембраны мышечного волокна так же, как и по мембране нервного волокна.
12
Безмиелиновые нервные волокна находятся преимущественно в составе автономной, или вегетативной, нервной системы. Нейролеммоциты оболочек безмиелиновых нервных волокон, располагаясь плотно, образуют тяжи. В нервных волокнах внутренних органов, как правило, в таком тяже имеется не один, а несколько осевых цилиндров, принадлежащих различным нейронам. Они могут, покидая одно волокно, переходить в соседнее. Такие волокна, содержащие несколько осевых цилиндров, называются волокнами кабельного типа. По мере погружения осевых цилиндров в тяж нейролеммоцитов оболочки последних прогибаются, плотно охватывают осевые цилиндры и, смыкаясь над ними, образуют глубокие складки, на дне которых и располагаются отдельные осевые цилиндры. Сближенные в области складки участки оболочки нейролеммоцита образуют сдвоенную мембрану — мезаксон, на которой как бы подвешен осевой цилиндр. Миелиновые нервные волокна встречаются как в центральной, так и в периферической нервной системе. Они значительно толще безмиелиновых нервных волокон. Они также состоят из осевого цилиндра, «одетого» оболочкой из нейролеммоцитов Шванна, но диаметр осевых цилиндров этого типа волокон значительно толще, а оболочка сложнее.
В процессе миелинизации аксон погружается в желобок на поверхности нейролеммоцита. Края желобка смыкаются. При этом образуется двойная складка плазмолеммы нейролеммоцита — мезаксон. Мезаксон удлиняется, концентрически наслаивается (как бы накручивается) на осевой цилиндр и образует вокруг него плотную слоистую зону — миелиновый слой. Отсутствие миелинового слоя в области узловых перехватов объясняется тем, что в этом участке волокна кончается один нейролеммоцит и начинается другой. Осевой цилиндр в этом месте частично прикрыт интердигитирующими отростками нейролеммоцитов. Оболочка аксона (аксолемма) обладает в области перехвата значительной электронной плотностью.
Миелиновые волокна центральной нервной системы не имеют насечек миелина, а нервные волокна не окружены базальными мембранами. Скорость передачи импульса миелиновыми волокнами больше, чем безмиелиновыми. Тонкие волокна, бедные миелином, и безмиелиновые волокна проводят нервный импульс со скоростью 1—2 м/с, тогда как толстые миелиновые — со скоростью 5—120 м/с. В безмиелиновом волокне волна деполяризации мембраны идет по всей аксолемме, не прерываясь, а в миелиновом возникает только в области перехватов. Таким образом, для миелиновых волокон характерно сальтаторное проведение возбуждения, т.е. прыжками. Между перехватами идет электрический ток, скорость которого выше, чем прохождение волны деполяризации по аксолемме.
К экстерорецепторам (внешним) относятся: слуховые, зрительные, обонятельные, вкусовые и осязательные рецепторы. К интерорецепторам (внутренним) относятся: висцеро-рецепторы (сигнализирующие о состоянии внутренних органов) и проприорецепторы (или рецепторы опорно-двигательного аппарата). В зависимости от специфичности раздражения, воспринимаемого данным видом рецептора, все чувствительные окончания делят на механорецепторы, барорецепторы, хеморецепторы, терморецепторы и некоторые другие. По особенностям строения чувствительные окончания подразделяют на свободные нервные окончания, т.е. состоящие только из конечных ветвлений осевого цилиндра, и несвободные, содержащие в своем составе все компоненты нервного волокна, а именно ветвления осевого цилиндра и клетки глии. Несвободные окончания, кроме того, могут быть покрыты соединительнотканной капсулой, и тогда они называются инкапсулированными. Несвободные нервные окончания, не имеющие соединительнотканной капсулы, называются неинкапсулированными. Свободные нервные окончания обычно воспринимают холод, тепло и боль. Такие окончания характерны для эпителия. В этом случае миелиновые нервные волокна подходят к эпителиальному пласту, теряют миелин, а осевые цилиндры проникают в эпителий и распадаются там между клетками на тонкие терминальные ветви. Инкапсулированные рецепторы соединительной ткани при всем их разнообразии всегда состоят из ветвления осевого цилиндра и глиальных клеток. Снаружи такие рецепторы покрыты соединительнотканной капсулой. Примером подобных окончаний могут служить весьма распространенные у человека пластинчатые тельца (или тельца Фатера-Пачини). В центре такого тельца располагается внутренняя луковица, или колба, образованная видоизмененными леммоцитами. Миелиновое чувствительное нервное волокно теряет около пластинчатого тельца миелиновый слой, проникает во внутреннюю луковицу и разветвляется. Снаружи тельце окружено слоистой капсулой, состоящей из фибробластов и спирально ориентированных волокон. Заполненные жидкостью пространства между пластинками содержат коллагеновые микрофибриллы. Давление на капсулу передается через заполненные жидкостью пространства между пластинками на внутреннюю луковицу и воспринимается безмиелиновыми волокнами во внутренней луковице. Пластинчатые тельца воспринимают давление и вибрацию. Они присутствуют в глубоких слоях дермы (особенно в коже пальцев), в брыжейке и внутренних органах. К чувствительным инкапсулированным окончаниям относятся осязательные тельца — тельца Мейснера. Эти структуры имеют овоидную форму, располагаются в верхушках соединительнотканных сосочков кожи. Осязательные тельца состоят из видоизмененных нейролеммоцитов — тактильных клеток, расположенных перпендикулярно длинной оси тельца. Части тактильных клеток, содержащие ядра, расположены на периферии, а уплощенные части, обращенные к центру, формируют пластинчатые отростки, интердигитирующие с отростками противоположной стороны. Тельце окружено тонкой капсулой. Миелиновое нервное волокно входит в основание тельца снизу, теряет миелиновый слой и формирует ветви, извивающиеся между тактильными клетками. Коллагеновые микрофибриллы и волокна связывают тактильные клетки с капсулой, а капсулу с базальным слоем эпидермиса, так что любое смещение эпидермиса передается на осязательное тельце Мейснера. К инкапсулированным нервным окончаниям относятся также рецепторы мышц и сухожилий: это нервно-мышечные веретена и нервно-сухожильные веретена. Нервно-мышечные веретена являются сенсорными органами в скелетных мышцах, которые функционируют как рецептор на растяжение. Веретено состоит из нескольких исчерченных мышечных волокон — интрафузальных волокон, заключенных в растяжимую соединительнотканную капсулу. Между капсулой и интрафузальными волокнами имеется заполненное жидкостью пространство. Интрафузальные волокна имеют актиновые и миозиновые миофиламенты только на концах, которые и сокращаются. Рецепторной частью интрафузального мышечного волокна является центральная, несокращающаяся часть. К интрафузальным мышечным волокнам подходят афферентные нервные волокна. При расслаблении (или растяжении) мышцы увеличивается и длина интрафузальных волокон, что регистрируется рецепторами. Одни окончания реагируют на изменение длины мышечного волокна и на скорость этого изменения, другие — реагируют только на изменение длины. При внезапном растяжении в спинной мозг поступает сильный сигнал, вызывающий резкое сокращение мышцы, с которой поступил сигнал, — динамический рефлекс на растяжение. При медленном, длительном растяжении волокна возникает статический сигнал на растяжение. Этот сигнал может поддерживать мышцу в состоянии сокращения в течение нескольких часов. Интрафузальные волокна имеют также эфферентную иннервацию. К ним подходят тонкие моторные волокна, оканчивающиеся аксо-мышечными синапсами на концах мышечного волокна. Вызывая сокращение концевых участков интрафузального волокна, они усиливают растяжение центральной рецепторной его части, повышая реакцию рецептора. Нервно-сухожильные веретена обычно располагаются в месте соединения мышцы с сухожилием. Коллагеновые пучки сухожилия, связанные с 10—15 мышечными волокнами, окружены соединительнотканной капсулой. К нервно-сухожильному веретену подходит толстое миелиновое волокно, которое теряет миелин и образует терминали, ветвящиеся между пучками коллагеновых волокон сухожилия. Сигнал с нервно-сухожильных веретен, вызванный напряжением мышцы, возбуждает тормозные нейроны спинного мозга. Последние тормозят соответствующие двигательные нейроны, предотвращая перерастяжение мышцы.
14
В зависимости от локализации окончаний терминальных веточек аксона, межнейрональные синапсы различают: аксо-дендритические, аксо-соматические, аксо-аксональные. Химические синапсы передают импульс на другую клетку с помощью специальных биологически активных веществ — нейромедиаторов, или нейротрансмиттеров, находящихся в синаптических пузырьках. Терминаль аксона представляет собой пресинаптическую часть, а область второго нейрона, или другой иннервируемой клетки, с которой она контактирует, — постсинаптическую часть. В пресинаптической части находятся синаптические пузырьки, многочисленные митохондрии и отдельные нейрофиламенты. Форма и содержимое синаптических пузырьков связаны с функцией синапса. Если передача импульса совершается с помощью медиатора ацетилхолина, - синапсы называют холинергическими, если медиатором служит норадреналин - адренергическими. В зависимости от передаваемого сигнала, нейромедиаторы, и соответственно синапсы, могут быть возбуждающими или тормозными. Такие нейромедиаторы, как дофамин, глицин и гамма-аминомасляная кислота (ГАМК) являются медиаторами тормозящих синапсов. Область синаптического контакта между двумя нейронами состоит из пресинаптической мембраны, синаптической щели и постсинаптической мембраны. Пресинаптическая мембрана — это мембрана клетки, передающей импульс. В этой области локализованы кальциевые каналы, способствующие слиянию синаптических пузырьков с пресинаптической мембраной и выделению медиатора в синаптическую щель.
Постсинаптическая мембрана — это участок плазмолеммы клетки, воспринимающий медиаторы и генерирующий импульс. Она снабжена рецепторными зонами для восприятия соответствующего нейромедиатора.
15 это концевые аппараты аксонов двигательных клеток соматической или вегетативной нервной системы. При их участии нервный импульс передается на ткани рабочих органов. Двигательные окончания в поперечнополосатых мышцах называются нервно-мышечными окончаниями. Они представляют собой окончания аксонов клеток двигательных ядер передних рогов спинного мозга или моторных ядер головного мозга. Нервно-мышечное окончание состоит из концевого ветвления осевого цилиндра нервного волокна и специализированного участка мышечного волокна. Миелиновое нервное волокно, подойдя к мышечному волокну, теряет миелиновый слой и погружается в него, вовлекая за собой его плазмолемму и базальную мембрану. Мембрана мышечного волокна образует многочисленные складки, формирующие вторичные синаптические щели эффекторного окончания. В области окончания мышечное волокно не имеет типичной поперечной исчерченности и характеризуется обилием митохондрий, скоплением круглых или слегка овальных ядер. Терминальные ветви нервного волокна в нервно-мышечном соединении характеризуются обилием митохондрий и многочисленными пресинаптическими пузырьками, содержащими характерный для этого вида окончаний медиатор — ацетилхолин. При возбуждении ацетилхолин поступает через пресинаптическую мембрану в синаптическую щель на холинорецепторы постсинаптической (мышечной) мембраны, вызывая ее возбуждение (волну деполяризации). Постсинаптическая мембрана моторного нервного окончания содержит фермент ацетилхолинэстеразу, разрушающий медиатор и ограничивающий этим срок его действия. Двигательные нервные окончания в гладкой мышечной ткани представляют собой чёткообразные утолщения (или варикозы) нервного волокна, идущего среди неисчерченных гладких миоцитов. Варикозы содержат адренергические или холинергические пресинаптические пузырьки. Нейролеммоциты в области варикозов часто отсутствуют, и волокно проходит «обнаженным». Сходное строение имеют секреторные нервные окончания (нейрожелезистые). Они представляют собой концевые утолщения терминали или утолщения по ходу нервного волокна, содержащие пресинаптические пузырьки, главным образом холинергические.
|
|||||||||
|
|
Последнее изменение этой страницы: 2018-06-01; просмотров: 365. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |
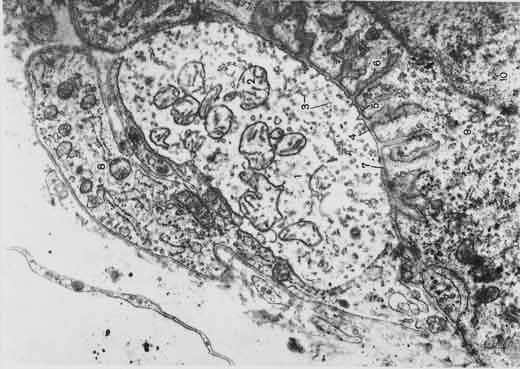
 Основная биологическая роль молекулы гистосовместимости 1-го класса — дать возможность Т-лимфоцитам с помощью своих Т-клеточных рецепторов распознать "свои и чужие" клетки. Информацию об этом несет разнообразный по составу и последовательности расположения аминокислот низкомолекулярный белок — эпитоп. Это фрагмент, возникший из белков, присутствующих внутри клетки. Среди них могут быть: а) фрагменты собственных белков, тогда они не вызывают ответной реакции со стороны специализированных Т-лимфоцитов; б) молекулы белка, кодируемые вирусом; в) молекулы белка, кодируемые мутантным геномом раковой клетки организма. В случае появления двух последних происходит реакция клетки, несущей чужеродный эпитоп, с Т-лимфоцитом, который называется цитотоксическим.
Основная биологическая роль молекулы гистосовместимости 1-го класса — дать возможность Т-лимфоцитам с помощью своих Т-клеточных рецепторов распознать "свои и чужие" клетки. Информацию об этом несет разнообразный по составу и последовательности расположения аминокислот низкомолекулярный белок — эпитоп. Это фрагмент, возникший из белков, присутствующих внутри клетки. Среди них могут быть: а) фрагменты собственных белков, тогда они не вызывают ответной реакции со стороны специализированных Т-лимфоцитов; б) молекулы белка, кодируемые вирусом; в) молекулы белка, кодируемые мутантным геномом раковой клетки организма. В случае появления двух последних происходит реакция клетки, несущей чужеродный эпитоп, с Т-лимфоцитом, который называется цитотоксическим.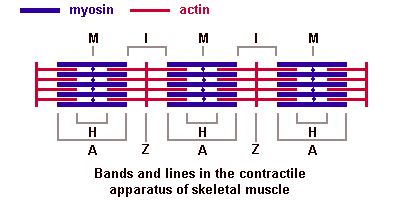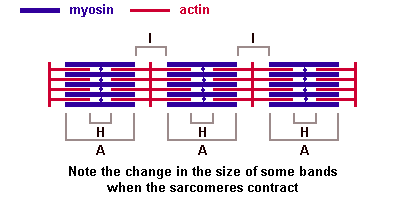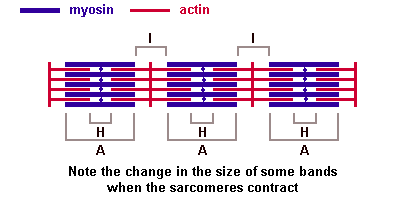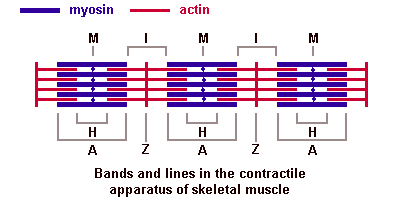 трубочка контактирует с двумя терминальными цистернами саркоплазматической сети, что обеспечивает выброс ионов кальция, необходимых для сократительного акта. В области Z-линий дискаIсходятся концы актиновых миофиламентов. Миозиновые миофиламенты, содержащие белок миозин, образуют анизотропные (А) темные диски, обладающие двойным лучепреломлением. В центре диска А проходит М-линия — мезофрагма. В М-линни сходятся концы миозиновых миофибрилл и обнаружена сеть канальцев саркоплазматической сети. Чередование в миофибриллах темных и светлых дисков придает мышечному волокну поперечную исчерченность. Структурной единицей миофибрилл является миомер (саркомер) — это участок миофибриллы между двумя Z-линиями. Его формула — А+21/2I.
трубочка контактирует с двумя терминальными цистернами саркоплазматической сети, что обеспечивает выброс ионов кальция, необходимых для сократительного акта. В области Z-линий дискаIсходятся концы актиновых миофиламентов. Миозиновые миофиламенты, содержащие белок миозин, образуют анизотропные (А) темные диски, обладающие двойным лучепреломлением. В центре диска А проходит М-линия — мезофрагма. В М-линни сходятся концы миозиновых миофибрилл и обнаружена сеть канальцев саркоплазматической сети. Чередование в миофибриллах темных и светлых дисков придает мышечному волокну поперечную исчерченность. Структурной единицей миофибрилл является миомер (саркомер) — это участок миофибриллы между двумя Z-линиями. Его формула — А+21/2I.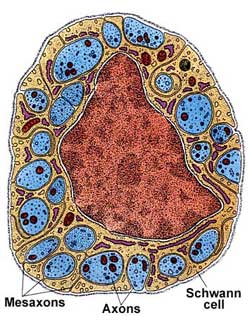 В центральной нервной системе оболочки отростков нейронов образуются отростками олигодендроглиоцитов, а в периферической — нейролеммоцитами Шванна.
В центральной нервной системе оболочки отростков нейронов образуются отростками олигодендроглиоцитов, а в периферической — нейролеммоцитами Шванна. Миелиновый слой оболочки такого волокна содержит значительное количество липидов, поэтому при обработке осмиевой кислотой он окрашивается в темно-коричневый цвет. В миелиновом слое периодически встречаются узкие светлые линии—насечки миелина, или насечки Шмидта — Лантермана. Через определенные интервалы (1—2 мм) видны участки волокна, лишенные миелинового слоя, — это т.н. узловатые перехваты, или перехваты Ранвье.
Миелиновый слой оболочки такого волокна содержит значительное количество липидов, поэтому при обработке осмиевой кислотой он окрашивается в темно-коричневый цвет. В миелиновом слое периодически встречаются узкие светлые линии—насечки миелина, или насечки Шмидта — Лантермана. Через определенные интервалы (1—2 мм) видны участки волокна, лишенные миелинового слоя, — это т.н. узловатые перехваты, или перехваты Ранвье. Отрезок волокна между смежными перехватами называется межузловым сегментом. Длина межузлового сегмента, так же как и толщина миелинового слоя, зависит от толщины осевого цилиндра. Насечка миелина (Шмидта—Лантермана) представляет собой участок миелинового слоя, где завитки мезаксона лежат неплотно друг к другу, образуя спиральный туннель, идущий снаружи внутрь и заполненный цитоплазмой нейролеммоцита, т.е. место расслоения миелина. Снаружи от нейролеммоцита располагается базальная мембрана.
Отрезок волокна между смежными перехватами называется межузловым сегментом. Длина межузлового сегмента, так же как и толщина миелинового слоя, зависит от толщины осевого цилиндра. Насечка миелина (Шмидта—Лантермана) представляет собой участок миелинового слоя, где завитки мезаксона лежат неплотно друг к другу, образуя спиральный туннель, идущий снаружи внутрь и заполненный цитоплазмой нейролеммоцита, т.е. место расслоения миелина. Снаружи от нейролеммоцита располагается базальная мембрана. Эти нервные окончания — рецепторы — рассеяны по всему организму и воспринимают различные раздражения как из внешней среды, так и от внутренних органов. Соответственно выделяют две большие группы рецепторов: экстерорецепторы и интерорецепторы.
Эти нервные окончания — рецепторы — рассеяны по всему организму и воспринимают различные раздражения как из внешней среды, так и от внутренних органов. Соответственно выделяют две большие группы рецепторов: экстерорецепторы и интерорецепторы. это структуры, предназначенные для передачи импульса с одного нейрона на другой или на мышечные и железистые структуры. Синапсы определяют направление проведения импульса. Если раздражать аксон электрическим током, импульс пойдет в обоих направлениях; но импульс, идущий в сторону тела нейрона и его дендритов, не может быть передан на другие нейроны. Только импульс, достигающий терминалей аксона, с помощью синапсов может передать возбуждение на другой нейрон, мышечную или железистую клетку. В зависимости от способа передачи импульса синапсы могут быть химическими или электрическими (электротоническими).
это структуры, предназначенные для передачи импульса с одного нейрона на другой или на мышечные и железистые структуры. Синапсы определяют направление проведения импульса. Если раздражать аксон электрическим током, импульс пойдет в обоих направлениях; но импульс, идущий в сторону тела нейрона и его дендритов, не может быть передан на другие нейроны. Только импульс, достигающий терминалей аксона, с помощью синапсов может передать возбуждение на другой нейрон, мышечную или железистую клетку. В зависимости от способа передачи импульса синапсы могут быть химическими или электрическими (электротоническими).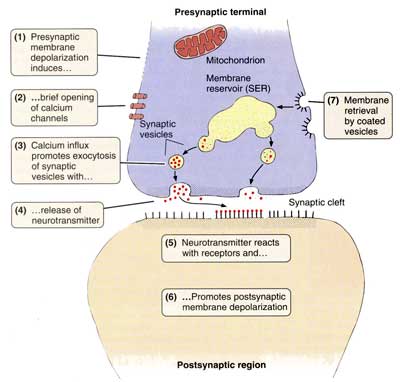 Синаптическая щель между пре- и постсинаптической мембранами имеет ширину 20—30 нм. Мембраны прочно прикреплены друг к другу в синаптической области филаментами, пересекающими синаптическую щель.
Синаптическая щель между пре- и постсинаптической мембранами имеет ширину 20—30 нм. Мембраны прочно прикреплены друг к другу в синаптической области филаментами, пересекающими синаптическую щель.