|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
Факторы, влияющие на метаболизм ксенобиотиковСпособность органов и тканей метаболизировать ксенобиотики зависит от набора и активности энзимов, участвующих в процессе. В значительной степени активность энзимов является внутренней характеристикой конкретной ткани, определяется генетическими особенностями организма и зависит от пола и возраста. Дополнительными факторами, порой существенным образом влияющими на содержание и активность энзимов, являются условия окружающей среды. Это прежде всего химические вещества, выступающие в качестве индукторов или ингибиторов энзимов, питание и действие патогенных факторов (таблица 5). Таблица 5. Факторы, влияющие на метаболизм ксенобиотиков
Генетические факторы Особи одного и того же вида живых существ порой значительно различаются по способности метаболизировать ксенобиотики. Это во многом детерминировано генетически. Так, в популяции людей выявляются лица, обладающие пониженной активностью цитохромР-450 зависимых оксидаз. "Слабые метаболизанты" могут отличаться отсутствием некоторых изоэнзимов, необходимых для катализа ряда превращений ксенобиотиков. Полиморфизм метаболизма ксенобиотиков отмечен для процессов ацетилирования ароматических веществ, содержащих амино-, сульфо-, амидную группу. Лица со слабым напряжением процессов ацетилирования более подвержены некоторым аллергическим реакциям, вызываемым химическими веществами. С другой стороны, излишне напряженный процесс ацетилирования ксенобиотиков может иметь пагубные последствия в результате биоактивации арилгидразинов.  Пол и возраст В опытах на лабораторных животных, в основном грызунах, показано, что половые гормоны принимают участие в регуляции активности энзимов метаболизма ксенобиотиков и прежде всего монооксигеназ. Так, взрослые самцы крыс быстрее метаболизируют такие вещества, как гексабарбитал, аминопирин, аминофенол и т.д. Вместе с тем анилин и его аналоги подвергаются биопревращению в организме самцов и самок с одинаковой скоростью. Кастрация нередко сопровождается снижением скорость метаболизма ксенобиотиков. У человека половые различия выражены не столь существенно. Отличия метаболизма ксенобиотиков, обусловленные возрастом, наиболее отчетливо проявляются у новорожденных и лиц пожилого возраста. Хорошо известно, что недостаточно развитая система метаболизма ксенобиотиков у новорожденных делает их особенно чувствительными к ряду токсикантов. Токсический процесс может стать следствием как накапливающихся в организме исходных продуктов, так и промежуточных метаболитов, не подвергающихся дальнейшей биотрансформации. В старческом возрасте наблюдается снижение клиаренса ксенобиотиков отчасти обусловленное понижением интенсивности метаболизма. Нарушение метаболизма ксенобиотиков печенью в старческом возрасте может являться следствием уменьшения интенсивности печеночного кровотока, хронических патологических процессов в печени, связанного с возрастом снижения активности ферментов. Влияние химических веществ Ксенобиотики, поступающие в организм, могут оказывать влияние на процессы метаболизма как самих этих веществ, так и других соединений, поступающих в организм одновременно или вслед за ними. Теоретически можно выделить три группы химических соединений, по-разному влияющих на метаболизм чужеродных веществ: 1. Практически не влияющие на активность энзимов метаболизма; 2. Повышающие активность энзимов - индукторы; 3. Угнетающие активность энзимов - ингибиторы. Важно иметь в виду, что одно и то же вещество может выступать и как индуктор и как ингибитор метаболизма другого вещества, в зависимости от того в каком порядке ксенобиотики поступают в организм - сукцессии или комбинации (см. ниже). Индукция энзимов Многие химические вещества, как эндогенные, так и поступающие из окружающей среды, обладают способностью усиливать синтез в организме энзимов биотрансформации ксенобиотиков. Этот феномен, получивший название индукции энзимов, существенным образом определяет чувствительность живых существ к действию токсикантов. Несколько сот химических веществ совершенно разного строения, как установлено, являются индукторами монооксигеназ и других ферментативных систем. К числу сильных индукторов микросомальных ферментов принадлежат многие лекарства и промышленные токсиканты. Все индукторы - жирорастворимые органические вещества. Их действие, как правило, неспецифично, то есть индуктор вызывает повышение активности более чем одного энзима. Индукция возможна, как правило, при повторном введении соединения. Индукторы метаболизма Многочисленные индукторы монооксигеназных систем можно отнести к одному из двух классов. Представителем первого класса является фенобарбитал, другие барбитураты, некоторые лекарства и инсектициды. Ко второму классу индукторов относятся в основном полициклические углеводороды: ТХДД, 3-метилхолантрен, бенз(а)пирен и т.д. Самым сильным из известных индукторов монооксигеназ является 2,3,7,8-тетрахлордибензо-р-диоксин (ТХДД). Его эффективная доза составляет 1 мкг/кг массы. В подавляющем большинстве случаев ксенобиотики проявляют свойства индукторов, действуя в значительно больших дозах (более 10 мг/кг). Фенобарбитал вызывает выраженную пролиферацию гладкого эндоплазматического ретикулума в гепатоцитах и увеличение активности Р-450. В результате возрастает мощность таких процессов, как деметилирование ксенобиотиков (нитроанизол), гидроксилирование (барбитураты), эпоксидирование (альдрин). Индукция, вызываемая полициклическими углеводородами не сопровождается пролиферацией гладкого эндоплазматического ретикулума, но при этом существенно возрастает активность Р-450, УДФГ-трансферазы, гидроксиолаз. Некоторые индукторы способны специфично активировать отдельные изоформы Р-450. К числу таковых относятся, в частности, прегненолол-16 Поскольку ксенобиотики, как правило, вызывают индукцию более чем одной ферментативной системы (барбитураты, полигалогенированные бифенилы одновременно вызывают индукцию Р-450, УДФГТ, GST и др.), предсказать влияние индукторов на токсикокинетику и токсикодинамику ксенобиотика практически не возможно. Эффект может быть определен только экспериментально. Задача усложняется еще и тем, что индукция того или иного энзима, вызванная разными индукторами, не одинаково сказывается на скорости метаболизма разных ксенобиотиков (таблица 6). Таблица 6. Влияние некоторых индукторов метаболизма на активность УДФ-глюкуронилтрансферазы печени крыс (при использовании
(K.W. Bock, 1977) Механизмы индукции Индукция предполагает синтез дополнительного количества того или иного энзима в органах и тканях de novo. Ингибиторы синтеза белка (пуромицин, этионин, циклогексимид), а также ингибиторы синтеза РНК (актиномицин Д) блокируют индукцию микросомальных энзимов. Поскольку блокаторы синтеза ДНК (гидроксимочевина) не эффективны, можно сделать вывод, что феномен индукции энзимов биотрансформации ксенобиотиков реализуется на уровне транскрипции генетической информации. Механизм феномена полностью не изучен. Одна из первых гипотез была предложена S. Granick в 1966 году для объяснения индукторных свойств стероидов. Автор полагал, что индукция эндоплазматического ретикулума гепатоцитов, содержащего цитохромы, регулируется концентрацией в клетке свободного гема, высвобождающегося в ходе обменных процессов в эндоплазматическом ретикулуме. Гем взаимодействует с внутриядерным апорепрессором. При этом образуется репрессор, который в свою очередь угнетает ген-оператор. В тот момент, когда ген-оператор находится в неактивной форме, на соответствующем участке ДНК не осуществляется синтез мРНК, необходимой для синтеза энзиматических белков, входящих в структуру эндоплазматического ретикулума. При отсутствии свободного гема или в тех случаях, когда гем не может присоединиться к апорепрессору, происходит активация синтеза мРНК и выраженная пролиферация эндоплазматического ретикулума. Естественными ингибиторами образования репрессора и являются стероиды, активно взаимодействующие с апорепрессором. В соответствии с этой моделью около 300 ксенобиотиков (далеко не все структурные аналоги стероидов) способны избирательно взаимодействовать с апорепрессором и блокировать тем самым образование репрессора. Эти вещества проявляют свойства индукторов микросомальных ферментов. Кроме того, так как большинство индукторов являются, как и стероиды, жирорастворимыми веществами они активно захватываются мембранными структурами цитоплазмы гепатоцитов, вытесняя при этом из соответствующих сайтов связывания "депонированные" стероиды. Концентрация последних повышается в цитоплазме, а затем и в ядре клеток. Связывание с апорепрессором усиливается, активируется процесс синтеза энзимов метаболизма ксенобиотиков. в соответствие с более поздними представлениями механизм действия стероидных гормонов, полициклических ароматических углеводородов, 2,3,7,8,-тетрахлордибензо-п-диоксина, состоит во взаимодействии с цитозольными рецепторными белками. Образующиеся комплексы мигрирует в ядро клетки, где вызывают дерепрессию регуляторных генов и, тем самым, активирует синтез того или иного энзима. В случае ТХДД такой рецепторный цитоплазматический протеин идентифицирован, по крайней мере, в гепатоцитах лини мышей, чувствительных к ароматическим углеводородам. Установлено, что синтез гидроксилазы ароматических улеводородов (aryl hydrocarbon hydroxylase) регулируется локусом единственного доминантного гена, Ah, и может быть усилен при введении ТХДД. Цитозольный белок-регулятор гена получил название Ah-рецепторный протеин. Другие индукторы, такие как барбитураты, вероятно действуют с помощью иного механизма. Хотя известно, что в основе процесса также лежит индукция синтеза белка, до конца не выяснено каким образом клетка распознает индуктор и как осуществляется воздействие на процесс транскрипции. Индукторы класса фенобарбитала относятся к числу малоактивных соединений. Для реализации эффекта нужны дозы препаратов на несколько порядков превышающие эффективные дозы ТХДД. Рецепторный белок для фенобарбитала пока не идентифицирован. Помимо усиления синтеза энзимов дополнительным механизмом индукции является стабилизация информационной РНК и белковых молекул в клетке. |
||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2018-04-12; просмотров: 392. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |
||||||||||||||||||||||||||
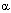 -карбонитрил (ПКН), активирующий 3А1 изоформу Р-450, этанол, индуцирующий 1А2 изоформу Р-450, клофибрат - 4А изоформу.
-карбонитрил (ПКН), активирующий 3А1 изоформу Р-450, этанол, индуцирующий 1А2 изоформу Р-450, клофибрат - 4А изоформу.