|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
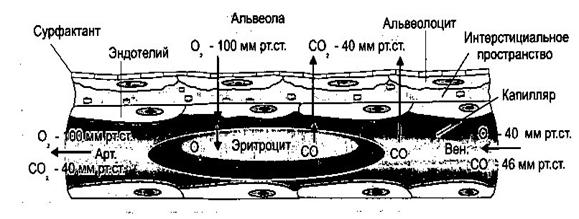
Величины парциальных давлений газов в атмосферном и альвеолярном воздухе
Парциальное давление кислорода в альвеолах (100 мм рт.ст.) значительно выше, чем напряжение кислорода в венозной крови, поступающей в капилляры легких (40 мм рт.ст.). Градиент парциального давления углекислого газа направлен в обратную сторону (46 мм рт.ст. в начале легочных капилляров и 40 мм рт.ст. в альвеолах). Эти градиенты давлений являются движущей силой диффузии кислорода и углекислого газа в крови и в альвеолярном воздухе. Молекулы газа в силу диффузии переходят из области большего парциального давления в область низкого парциального давления. На рис.11. приведены величины парциального давления и напряжения газов в альвеолярном воздухе, артериальной и венозной части легочного капилляра, определяющие процессы диффузии. Проницаемость мембраны для газа выражают величиной диффузионной способности легких – это количество газа, проникающего через легочную мембрану за 1 мин. при градиенте давления в 1 мм рт.ст. В норме диффузионная способность для О2 составляет 20-25 мл/мин/мм рт.ст. Коэффициент диффузии для СО2 в 20-25 раз больше, чем для кислорода (диффузионная способность для СО2 составляет 600 мл на 1 мм рт.ст. в минуту).
Рис.11. Диффузия газов между альвеолярным воздухом и кровью легочного капилляра, на основе разницы парциальных давлений и парциальных напряжений. При прочих равных условиях углекислый газ диффундирует через определенный слой среды в 20-25 раз быстрее, чем кислород. Именно поэтому обмен СО2 в легких происходит достаточно полно, несмотря на небольшой градиент парциального давления этого газа.  Транспорт газов кровью Кислород и углекислый газ в крови находятся в двух состояниях: в химически связанном и в растворенном. Хотя содержание в крови кислорода и углекислого газа в физически растворенном состоянии относительно невелико, это состояние играет существенную роль в жизнедеятельности организма. Для того, чтобы связаться с теми или иными веществами, дыхательные газы сначала должны быть доставлены к ним в физически растворенном виде. Таким образом, при диффузии газов каждая молекула О2 и СО2 определенное время пребывает в состоянии физического растворения.
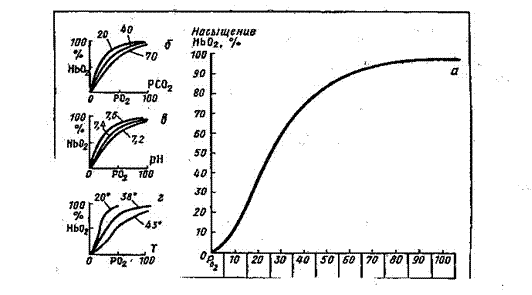
Рис.12. График диссоциации оксигемоглобина. а - при нормальном парциальном давлении СО2, б - влияние изменений парциального давления СО2, в – влияние изменений рН, г – влияние изменений температуры.
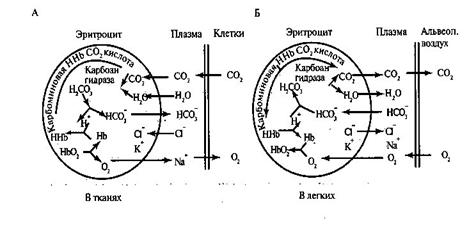
Транспорт кислорода. Большая часть кислорода переносится кровью в виде химического соединения с гемоглобином, образуя оксигемоглобин. Присоединение кислорода к гемоглобину (оксигенация гемоглобина) происходит без изменения валентности железа. Степень насыщения гемоглобина кислородом, т.е. образование оксигемоглобина, зависит от напряжения кислорода в крови. Графически эту зависимость отражает так называемая кривая диссоциации оксигемоглобина. Эта кривая имеет S – образную форму. Крутая часть графика диссоциации оксигемоглобина соответствует напряжению кислорода в тканях. Здесь усиливается диссоциация HbО2, и О2 из крови переходит в ткани. Отлогая правая часть графика соответствует высоким напряжениям кислорода и свидетельствует о том, что в этих условиях содержание оксигемоглобина мало зависит от напряжения кислорода и его парциального давления в альвеолярном воздухе. Эта часть графика соответствует процессам образования оксигемоглобина в легких. Сродство гемоглобина к кислороду изменяется в зависимости от многих факторов. Как правило, все эти факторы смещают кривую, увеличивая или уменьшая ее наклон, но не изменяя при этом ее S – образную форму. Если сродство гемоглобина к кислороду повышается, то процесс идет в сторону образования оксигемоглобина, и график диссоциации смещается влево. Это наблюдается при снижении напряжения углекислого газа при понижении температуры, при сдвиге рН в щелочную сторону (рис.12). При снижении сродства гемоглобина к кислороду процесс идет больше в сторону диссоциации оксигемоглобина, при этом график диссоциации смещается вправо. Это наблюдается при повышении парциального давления углекислого газа, при повышении температуры, при смещении рН в кислую сторону. При ряде патологических состояний наблюдаются изменения процесса транспорта кислорода кровью. Ряд заболеваний (например, некоторые виды анемий) сопровождается сдвигами кривой диссоциации оксигемоглобина вправо (реже – влево). Транспорт углекислого газа.Двуокись углерода – конечный продукт окислительных обменных процессов в клетках – переносится с кровью к легким в трех формах: в свободном, в виде кислых солей угольной кислоты и карбгемоглобина. Напряжение СО2 в артериальной крови, поступающей в тканевые капилляры, составляет 40 мм рт.ст. В клетках тканей наибольшее напряжение СО2 - 60 мм рт.ст., в тканевой жидкости 46 мм рт.ст. В связи с этим физически растворенный СО2 переносится по градиенту напряжения из тканевой жидкости в капилляры. В результате напряжение СО2 в венозной крови составит 46 мм рт.ст. Здесь большая часть СО2 претерпевает ряд химических превращений – происходит гидратация молекул СО2 с образованием угольной кислоты. В плазме эта реакция идет медленно, а в эритроцитах под влиянием фермента карбоангидразы она резко ускоряется примерно в 10 тыс. раз. Поскольку этот фермент присутствует только в клетках, практически все молекулы СО2, участвующие в реакции гидратации, должны сначала поступить в эритроциты. В зависимости от напряжения СО2 карбоангидраза катализирует как образование угольной кислоты, так и расщепление ее на двуокись углерода, и воду (в капиллярах легких). Таким образом, эритроциты переносят в растворенном виде в 3 раза больше СО2, чем плазма. Угольная кислота диссоциирует на ионы Н+ и НСО-3. Концентрация ионов НСО-3, образующихся при диссоциации солей угольной кислоты, в эритроцитах возрастает. Мембрана эритроцитов обладает высокой проницаемостью для анионов. Поэтому часть ионов НСО-3 поступает в плазму крови (рис.12). Взамен ионов НСО-3 в эритроциты из плазмы входят ионы Cl-, отрицательные заряды которых уравновешиваются ионами К+. В плазме крови увеличивается количество бикарбоната Na. Этот обменный процесс называется хлоридным сдвигом. Гемоглобин служит источником катионов, необходимых для связывания угольной кислоты в форме бикарбонатов. В эритроцитах тканевых капилляров образуется дополнительное количество бикарбоната К+, а также карбогемоглобина, а в плазме крови увеличивается количество бикарбоната Na+. В таком виде СО2 переносится к легким. СО2 может связываться также путем непосредственного присоединения к аминогруппам белкового компонента гемоглобина. При этом образуется так называемая карбаминова связь. Часть молекул СО2 соединяется в эритроцитах с гемоглобином, образуя карбгемоглобин. В капиллярах малого круга кровообращения напряжение СО2 снижается. От HbСО2 отщепляется СО2. Одновременно происходит образование HbО2. HbО2 вытесняет К+ из бикарбонатов. Угольная кислота в эритроцитах быстро разлагается на СО2 и Н2О. Ионы НСО3 входят в эритроциты, а ионы Cl- выходят в плазму крови, где уменьшается количество бикарбоната Na+. Двуокись углерода диффундирует в альвеолярный воздух (рис 13).
Рис.13. Динамика процессов, происходящих в плазме и эритроцитах при газообмене в тканях (А) и легких (Б). Содержание углекислого газа, находящегося в крови в виде химических соединений, непосредственно зависит от его напряжения и описывается кривой, аналогичной кривой диссоциации оксигемоглобина. Зависимость содержания СО2 от степени оксигенации гемоглобина называется эффектом Холдейна. Данный эффект частично обусловлен различной способностью оксигемоглобина и дезоксигемоглобина к образованию карбаминовой связи. Диффузия газов в тканях Парциальное напряжение СО2 в тканях равно 60 мм рт. ст. парциальное напряжение О2 в тканях снижается до нуля. Однако, в тканевой жидкости напряжение СО2 составляет 46 мм рт.ст., а кислорода 20 - 40 мм рт.ст. В артериальной крови, притекающей к тканям напряжение СО2 – меньше 40 мм рт.ст., а напряжение О2 чуть более 100 мм рт.ст. Происходит газообмен, О2 переходит в тканевую жидкость и в ткани, СО2 – в кровь. В результате кровь становится венозной и в ней напряжение СО2 равно 46 мм рт.ст., а напряжение О2 – 40 мм рт.ст. Кислород поступивший в ткани по градиенту парциального давления из крови тканевых капилляров достигает митохондрий. Переходу кислорода в ткани способствует раскрытие дополнительных капилляров, под влиянием продуктов обмена веществ, в частности гистамина. Тканевое дыхание – это механизмы доставки и потребления кислорода, а также образование и выделение СО2 из ткани. Путь кислорода охватывает три отрезка: транспорт кровью; диффузию из капилляров большого круга кровообращения в клетки и их митохондрии; химические реакции с цитохромной системой митохондрий для образования АТФ. На обмен кислорода и углекислого газа в тканях влияют площадь обменной поверхности, количество эритроцитов, протекающих по капиллярам в единицу времени, величина диффузионного расстояния и коэффициенты диффузии тех сред, через которые осуществляется перенос газов. III. РЕГУЛЯЦИЯ ДЫХАНИЯ Первоначально были изучены структуры мозга, которые непосредственно отвечают за акт вдоха и выдоха. Для этих целей производили перерезки и разрушения мозговых структур. Было установлено, что отделение головного мозга от спинного приводит к полной остановке дыхания. Следовательно, структуры, ответственные за процесс вдоха и выдоха, находятся в бульбопонтийной области мозга. А.Н. Миславский (1885 г.) обнаружил, что разрушение медиальной части продолговатого мозга в нижнем углу ромбовидной ямки приводит к полной остановке дыхания. Когда разрушают нейроны рострального отдела варолиевого моста, дыхательные циклы становятся редкими и нерегулярными. Люмсден (1923 г.) впервые наблюдавший это явление, сделал вывод, что в ростральных участках варолиевого моста находятся нейроны, которые ответственны за смену вдоха на выдох. Он назвал их пневмотаксическим центром, который способствует периодической деятельности дыхательного центра, увеличивает скорость развития инспираторной активности, повышает возбудимость выключения вдоха, ускоряет наступление следующей инспирации. При перерезках по верхнему краю варолиева моста дыхание становится ритмичным, с длинными паузами и задерживается на выдохе. Здесь находится апнейстический дыхательный центр. Гипоталамус и ретикулярная формация определяют участие дыхания в эмоциональных и вегетативных реакциях, а мозжечок контролирует акт дыхания при движении. В регуляции дыхания принимают участие и полушария большого мозга, которые участвуют в тонком адекватном приспособлении дыхания к меняющимся условиям существования организма. |
||||||||||||||
|
|
Последнее изменение этой страницы: 2018-04-12; просмотров: 451. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |