|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
Мышечная и суставная рецепция (проприорецепция)
В мышцах человека содержатся три типа специализированных рецепторов: первичные окончания веретён, вторичные окончания веретён и сухожильные рецепторы Гольджи. Эти рецепторы реагируют на механические раздражения и участвуют в координации движений, являясь источником информации о состоянии двигательного аппарата (см. также гл. 5). Мышечное веретено имеет длину в несколько миллиметров, ширину в десятые доли миллиметра, одето капсулой и расположено в толще мышцы. Внутри капсулы находится пучок интрафузальных мышечных волокон. Веретена расположены параллельно внешним по отношению к капсуле экстрафузальным волокнам, поэтому при растяжении мышцы нагрузка на веретена увеличивается, а при сокращении – уменьшается. В расслабленной мышце импульсация, идущая от веретён, невелика, но они реагируют повышением частоты разрядов на удлинение мышцы. Таким образом, веретена дают мозгу информацию о длине мышцы и её изменениях. Импульсация, идущая от веретён, в спинном мозге возбуждает мотонейроны своей мышцы и тормозит мотонейроны мышцы-антагониста, а также возбуждает мотонейроны сгибателей и тормозит мотонейроны разгибателей. Сухожильные рецепторы Гольджи находятся в зоне соединения мышечных волокон с сухожилием и расположены последовательно по отношению к мышечным волокнам. Они слабо реагируют на растяжение мышцы, но возбуждаются при её сокращении, причём их импульсация пропорциональна силе сокращения. Поэтому сухожильные рецепторы информируют мозг о силе, развиваемой мышцей. Идущие от этих рецепторов волокна в спинном мозге вызывают торможение мотонейронов собственной мышцы и возбуждение мотонейронов мышцы-антагониста. Информация от мышечных рецепторов по восходящим путям спинного мозга поступает в высшие отделы ЦНС, включая кору большого мозга.  Суставные рецепторы изучены меньше, чем мышечные. Известно, что они реагируют на положение сустава и на изменения суставного угла, участвуя таким образом в системе обратных связей от двигательного аппарата.
Передача и переработка соматосенсорной информации
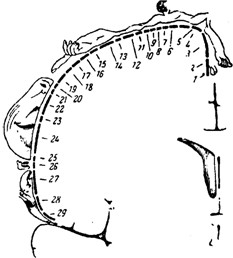
Чувствительность кожи и ощущение движения связаны с проведением в мозг сигналов от рецепторов по двум основным путям (трактам): лемнисковому и спиноталамическому, значительно различающимся по своим свойствам. Лемнисковый путь передаёт в мозг сигналы о прикосновении к коже, давлении на неё и движениях в суставах. Отличительная особенность этого пути – быстрая передача в мозг наиболее точной информации, дифференцированной по силе и месту воздействия. По мере перехода на всё более высокие уровни изменяются некоторые важные свойства нейронов лемнискового пути. Значительно увеличиваются рецептивные поля нейронов (в продолговатом мозге – в 2–30, а в коре большого мозга – в 15–100 раз). Несмотря на увеличение размеров рецептивных полей, нейроны остаются достаточно специфичными (нейроны поверхностного прикосновения, глубокого прикосновения, нейроны движения в суставах и нейроны положения или угла сгибания суставов). Для корковой части лемнискового пути характерна чёткая топографическая организация, т.е. проекция кожной поверхности осуществляется в центры мозга по принципу «точка в точку» [Кейдель, 1975; Сомьен, 1975; Дудел и др., 1985]. При этом площадь коркового представительства той или иной части тела определяется её функциональной значимостью: формируется так называемый «сенсорный гомункулюс» (рис. 4.10). Роль соматосенсорной коры состоит в интегральной оценке соматосенсорных сигналов, включении их в сферу сознания и в сенсорное обеспечение выработки новых двигательных навыков. Спиноталамический путь значительно отличается от лемнискового сравнительно медленной передачей афферентных сигналов, нечётко дифференцированной информацией о свойствах раздражителя и не очень чёткой её топографической локализацией; он служит для передачи температурной, всей болевой и в значительной мере тактильной чувствительности.
Рис. 4.10. Корковое представительство кожной чувствительности. Расположение в соматосенсорной зоне коры больших полушарий мозга человека проекций различных частей тела: 1– половые органы; 2 – пальцы ноги; 3 – ступня; 4 – голень; 5 – бедро; 6 – туловище; 7– шея; 8 – голова; 9 – плечо; 10–11 – локоть; 12 – предплечье; 13 – запястье; 14 – кисть; 15–19 – пальцы руки; 20 – глаза; 21 – нос; 22 – лицо; 23 – верхняя губа; 24,26 – зубы; 25 – нижняя губа; 27 – язык; 28 – глотка; 29 – внутренние органы. Размеры изображений частей тела соответствуют размерам их сенсорного представительства Болевая чувствительность практически не представлена на корковом уровне (раздражение коры не вызывает боли), поэтому считают, что высшим центром болевой чувствительности является таламус, где 60% нейронов в соответствующих ядрах чётко реагирует на болевое раздражение. Таким образом, спиноталамическая система играет важную роль в организации генерализованных ответов на действие болевых, температурных и тактильных раздражителей.
ОБОНЯТЕЛЬНАЯ СИСТЕМА
Рецепторы обонятельной системы расположены в области верхних носовых ходов. Обонятельный эпителий имеет толщину 100–150 мкм и содержит около 10 млн. рецепторных клеток диаметром 5–10 мкм, расположенных между опорными клетками (рис. 4.11). На поверхности каждой обонятельной клетки имеется сферическое утолщение – обонятельная булава, из которой выступает 6–12 волосков длиной до 10 мкм. Обонятельные волоски погружены в жидкую среду, вырабатываемую боуменовыми железами. Наличие подобных волосков в десятки раз увеличивает площадь контакта рецептора с молекулами пахучих веществ. От нижней части рецепторной клетки отходит аксон. Аксоны всех рецепторов образуют обонятельный нерв, который проходит через основание черепа и вступает в обонятельную луковицу. Молекулы пахучих веществ попадают в обонятельную слизь с постоянным током воздуха. Здесь они взаимодействуют с находящимся в волосках рецептора рецепторным белком. В результате этого взаимодействия в мембране рецептора открываются натриевые каналы, и генерируется рецепторный потенциал. Это приводит к импульсному разряду в аксоне рецептора – волокне обонятельного нерва. Каждая рецепторная клетка способна ответить возбуждением на характерный для неё, хотя и широкий, спектр пахучих веществ. Спектры чувствительности разных клеток сильно перекрываются. Вследствие этого более чем 50% пахучих веществ оказываются общими для любых двух обонятельных клеток. Электроольфактограммой называют суммарный электрический потенциал, регистрируемый от поверхности обонятельного эпителия. Это монофазная негативная волна длительностью в несколько секунд, возникающая даже при кратковременном воздействии пахучего вещества. Кодирование обонятельной информации. Одиночные рецепторы отвечают на запах увеличением частоты импульсации, которое зависит от качества и интенсивности стимула [Физиол. сенсорн. систем, 1972; Кейдель, 1975; Сомьен, 1975; Тамар, 1976; Батуев, Куликов, 1983; Дудел и др., 1985]. Каждый обонятельный рецептор отвечает на многие пахучие вещества, отдавая «предпочтение» некоторым из них. Считают, что на этих свойствах рецепторов может быть основано кодирование запахов и их опознание в центрах обонятельной системы. В обонятельной луковице регистрируется электрический ответ, который зависит от пахучего вещества: при разных запахах меняется пространственная мозаика возбуждённых и заторможенных участков обонятельной луковицы.
Рис. 4.11 . Схема строения обонятельного эпителия по данным электронной микроскопии: ОБ – обонятельная булава; ОК – опорная клетка; ЦО – центральные отростки обонятельных клеток; БК – базальная клетка; БМ – базальная мембрана; ВЛ – обонятельные волоски; МВР – микроворсинки обонятельных и МВО – микроворсинки опорных клеток Центральные проекции обонятельной системы. Выходящий из луковицы обонятельный тракт направляется в разные отделы мозга: переднее обонятельное ядро, обонятельный бугорок, препириформную кору, периамигдалярную кору и часть ядер миндалевидного комплекса. Обонятельная луковица связана также с гиппокампом, пириформной корой и другими отделами «обонятельного мозга» через несколько переключений. Наличие многих центров «обонятельного мозга» обеспечивает связь обонятельной системы с другими сенсорными системами и организацию на этой основе пищевого, оборонительного и полового поведения. Чувствительность обонятельной системы человека чрезвычайно велика: один обонятельный рецептор может быть возбуждён одной молекулой пахучего вещества, а возбуждение небольшого количества рецепторов приводит к возникновению ощущения. В то же время изменение интенсивности запаха (дифференциальный порог) оценивается людьми довольно грубо: наименьшее воспринимаемое различие в силе запаха составляет 30–60% от его исходной концентрации. Адаптация в обонятельной системе происходит сравнительно медленно (десятки секунд или минуты) и зависит от скорости потока воздуха над обонятельным эпителием и от концентрации пахучего вещества.
ВКУСОВАЯ СИСТЕМА
В процессе эволюции вкус формировался как механизм выбора или отвергания пищи. Выбор предпочитаемой пищи отчасти основан на врождённых механизмах, но в значительной мере зависит от связей, выработанных в онтогенезе. Вкус, так же как и обоняние, основан на хеморецепции и даёт информацию о характере и концентрации веществ, поступающих в рот. В результате запускаются реакции, изменяющие работу органов пищеварения или ведущие к удалению вредных веществ, попавших в рот. Вкусовые рецепторы сконцентрированы во вкусовых почках, расположенных на языке, задней стенке глотки, мягком нёбе, миндалине и надгортаннике. Больше всего их на кончике языка. Каждая из примерно 10 000 вкусовых почек человека состоит из нескольких рецепторных и опорных клеток. Вкусовая почка соединена с полостью рта через вкусовую пору. Вкусовая рецепторная клетка имеет длину 10–20 мкм и ширину 3–4 мкм и снабжена на конце, обращённом в просвет поры, 30–40 тончайшими микроворсинками. Считают, что они играют важную роль в рецепции химических веществ, адсорбированных в канале почки. Многие этапы преобразования химической энергии вкусовых веществ в энергию нервного возбуждения вкусовых рецепторов ещё неизвестны. Электрические потенциалы вкусовой системы. Суммарный потенциал рецепторных клеток возникает при раздражении языка сахаром, солью и кислотой. Он развивается медленно: максимум потенциала приходится на 10–15-й с после воздействия, хотя электрическая активность в волокнах вкусового нерва начинается раньше. Проводящие пути и центры вкуса. Проводниками для всех видов вкусовой чувствительности служат так называемая «барабанная струна» и языкоглоточный нерв, ядра которых расположены в продолговатом мозге. Многие из волокон специфичны, так как отвечают лишь на соль, кислоту, хинин или сахар. Наиболее убедительной считается гипотеза о том, что четыре основных вкусовых ощущения – горькое, сладкое, кислое и солёное – кодируются не импульсацией в одиночных волокнах, а распределением частоты разрядов в большой группе волокон, по-разному возбуждаемых вкусовым веществом. Афферентные сигналы, вызванные вкусовой стимуляцией, поступают в ядро одиночного пучка ствола мозга. От этого ядра аксоны вторых нейронов восходят в составе медиальной петли до таламуса, где расположены третьи нейроны, аксоны которых направляются в корковый центр вкуса.
|
||
|
|
Последнее изменение этой страницы: 2018-04-12; просмотров: 509. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |