|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
Строение и функции рецепторного вестибулярного аппарата
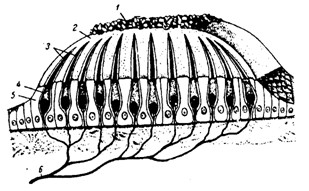
Периферическим отделом вестибулярной системы является вестибулярный аппарат, расположенный в пирамиде височной кости. Он состоит из преддверия и трёх полукружных каналов. Полукружные каналы (рис. 4.9) располагаются в трёх взаимно перпендикулярных плоскостях. Один из концов каждого канала расширен (ампула). Вестибулярный аппарат включает в себя также два мешочка. В них на возвышениях находится отолитовый аппарат: скопления рецепторных клеток. Выступающая в полость мешочка рецепторная клетка оканчивается длинным подвижным волоском и 60–80 склеенными неподвижными волосками. Они пронизывают желеобразную мембрану, содержащую кристаллики карбоната кальция – отолиты. Волосковые клетки возбуждаются при скольжении отолитовой мембраны по волоскам, т.е. при их сгибании. В перепончатых полукружных каналах, заполненных эндолимфой, рецепторные волосковые клетки сконцентрированы в ампулах. Во время угловых ускорений эндолимфа приходит в движение, волоски сгибаются и волосковые клетки возбуждаются. При противоположно направленном движении они тормозятся. Это связано с тем, что отклонение волоска в одну сторону приводит к открыванию каналов и деполяризации волосковой клетки, а отклонение в противоположном направлении закрывает каналы и гиперполяризует рецептор. В волосковых клетках преддверия и ампулы при их сгибании генерируется рецепторный потенциал, который усиливает выделение ацетилхолина и через синапсы активирует окончания волокон вестибулярного нерва. Волокна вестибулярного нерва (отростки биполярных нейронов) направляются в продолговатый

Рис. 4.9. Строение отолитового аппарата: 7 – отолиты; 2 – отолитовая мембрана; 3 – волоски рецепторных клеток; 4 рецепторные клетки; 5 – опорные клетки; 6 – нервные волокна. Продолговатый мозг. Импульсы, приходящие по этим волокнам, активируют нейроны бульбарного вестибулярного комплекса (ядра: преддверное верхнее Бехтерева, преддверное латеральное Дейтерса, Швальбе и др.). Отсюда сигналы направляются во многие отделы ЦНС: спинной мозг, мозжечок, глазодвигательные ядра, кору мозга, ретикулярную формацию и вегетативные ганглии.
Электрические явления в вестибулярной системе
Даже в полном покое в волокнах вестибулярного нерва регистрируется спонтанная импульсация. Частота разрядов в нерве повышается при поворотах головы в одну сторону и тормозится при поворотах в другую сторону (детекция направления движения). Две трети волокон обнаруживают эффект адаптации (уменьшение частоты разрядов) во время длящегося действия углового ускорения. Нейроны вестибулярных ядер обладают способностью реагировать и на изменение положения конечностей, повороты тела, сигналы от внутренних органов, т.е. осуществлять синтез информации, поступающей из разных источников. Нейроны вестибулярных ядер обеспечивают контроль и управление различными двигательными реакциями. Вестибуло-спинальные влияния изменяют импульсацию нейронов сегментарных уровней спинного мозга. Так осуществляется динамическое перераспределение тонуса скелетной мускулатуры и включаются рефлекторные реакции, необходимые для сохранения равновесия. В вестибуло-вегетативные реакции вовлекаются сердечно-сосудистая система, желудочно-кишечный тракт и другие внутренние органы. При сильных и длительных нагрузках на вестибулярный аппарат возникает болезнь движения (например, морская болезнь). Вестибуло-глазодвигательные рефлексы (глазной нистагм) состоят в медленном ритмическом движении глаз в противоположную вращению сторону, сменяющемся их скачком обратно. Возникновение и характеристики вращательного глазного нистагма – важные показатели состояния вестибулярной системы и широко используются в эксперименте и клинике.
Основные афферентные пути и проекции вестибулярных сигналов
Два основных пути поступления вестибулярных сигналов в кору мозга обезьян следующие: прямой – через вентральное постлатеральное ядро и непрямой – через вентролатеральное ядро. В коре основные афферентные проекции вестибулярного аппарата локализованы в задней части постцентральной извилины. В моторной коре кпереди и книзу от центральной борозды обнаружена вторая вестибулярная зона. Локализация вестибулярной зоны в коре мозга человека окончательно не выяснена.
Функции вестибулярной системы
Вестибулярная система помогает ориентироваться в пространстве при активном и пассивном движении. При пассивном движении лабиринтный аппарат с помощью корковых отделов системы анализирует и запоминает направление движения и повороты. Следует подчеркнуть, что в нормальных условиях пространственная ориентировка обеспечивается совместной деятельностью зрительной и вестибулярной систем. Чувствительность вестибулярной системы здорового человека очень высока: отолитовый аппарат позволяет воспринять ускорение прямолинейного движения, равное всего 2 см/с 2 . Порог различения наклона головы в сторону – всего около 1 углового градуса, а вперёд и назад – 1,5–2 угловых градуса. Рецепторная система полукружных каналов позволяет человеку замечать ускорения вращения в 2–3 угловых градуса в 1 с 2 .
СОМАТОСЕНСОРНАЯ СИСТЕМА
В соматосенсорную систему входят система кожной чувствительности и чувствительная система скелетно-мышечного аппарата, главная роль в которой принадлежит проприорецепции.
Кожная рецепция
Кожные рецепторы сосредоточены на огромной кожной поверхности (1,4–2,1 м 2 ). В коже находится множество рецепторов, чувствительных к прикосновению, давлению, вибрации, теплу и холоду, а также к болевым раздражениям. Они весьма различны по строению, локализуются на разной глубине кожи и распределены неравномерно по её поверхности. Больше всего их в коже пальцев рук, ладоней, подошв, губ и половых органов. У человека в коже с волосяным покровом (90% всей кожной поверхности) основным типом рецепторов являются свободные окончания нервных волокон, идущих вдоль мелких сосудов, а также более глубоко локализованные разветвления тонких нервных волокон, оплетающих волосяную сумку. Эти окончания обеспечивают высокую чувствительность волос к прикосновению. Рецепторами прикосновения являются также осязательные мениски (диски Меркеля), образованные в нижней части эпидермиса контактом свободных нервных окончаний с модифицированными эпителиальными структурами. Их особенно много в коже пальцев рук. В коже, лишённой волосяного покрова, находят много осязательных телец (телец Мейснера). Они локализованы в сосочковом слое кожи пальцев рук и ног, ладонях, подошвах, губах, языке, половых органах и сосках груди. Другими инкапсулированными нервными окончаниями, но более глубоко расположенными, являются пластинчатые тельца, или тельца Пачини (рецепторы давления и вибрации). Они имеются также в сухожилиях, связках, брыжейке. Теории кожной чувствительности многочисленны и во многом противоречивы. Наиболее распространено представление о наличии специфических рецепторов для четырёх основных видов кожной чувствительности: тактильной, тепловой, холодовой и болевой. Исследования электрической активности одиночных нервных окончаний и волокон свидетельствуют о том, что многие из них воспринимают лишь механические или температурные стимулы. Механизмы возбуждения кожных рецепторов. Механический стимул приводит к деформации мембраны рецептора. В результате этого электрическое сопротивление мембраны уменьшается, т.е. увеличивается её проницаемость для ионов. Через мембрану рецептора начинает течь ионный ток, приводящий к генерации рецепторного потенциала. При достижении рецепторным потенциалом критического уровня деполяризации генерируются импульсы, распространяющиеся по волокну в ЦНС. Адаптация кожных рецепторов. По скорости адаптации при длящемся действии раздражителя большинство кожных рецепторов подразделяются на быстрои медленноадаптирующиеся. Наиболее быстро адаптируются тактильные рецепторы, расположенные в волосяных фолликулах, а также пластинчатые тельца. Адаптация кожных механорецепторов приводит к тому, что мы перестаём ощущать постоянное давление одежды или привыкаем носить на роговице глаз контактные линзы.
|
||
|
|
Последнее изменение этой страницы: 2018-04-12; просмотров: 529. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |