|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
Характеристика мембранных белков
По выполняемым функциям белки в составе мембран делятся на 1. структурные; 2. каталитические; 3. рецепторные; 4. транспортные. Количество белков в мембранах могут существенно отличаться. Например, в миелиновой мембране, предназначенной для изоляции нервных волокон, белки составляют только 25% массы мембраны, а в мембранах митохондрий, связанных с процессами окислительного фосфорилирования, на долю белков приходится около 75% массы. В плазматической мембране доля белков и липидов примерно одинаковы. Углеводы в составе мембран не представлены самостоятельными соединениями, а обнаруживаются только в соединении с белками (гликопротеины) или липидами (гликолипиды). Длина углеводных цепей колеблется от двух до восемнадцати остатков моносахаридов. Большая часть углеводов расположена на наружной поверхности плазматической мембраны. Функции углеводов в биомембранах – контроль за межклеточными взаимодействиями, поддержание иммунного статуса, рецепция, обеспечение стабильности белковых молекул в мембране.  49. Липиды-амфипаты: представители, способность к агрегации, образованию мицелл, бислоев. Полярные липиды как компоненты биомембран и липопротеинов. Как уже упоминалось (9.1), компонентами липидов являются остатки жирных кислот и одно- или многоатомных спиртов. Примеры жирных кислот, встречающихся в составе липидов мембран, представлены на рисунке 9.4. Выучите эти формулы.
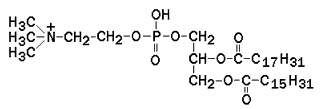
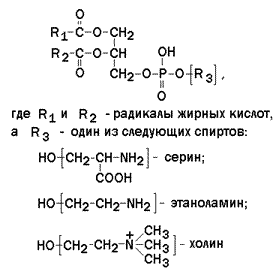
Рисунок 9.4. Наиболее часто встречающиеся природные жирные кислоты. Основные особенности строения жирных кислот, входящих в состав природных жиров: · они содержат чётное число атомов углерода (С16 – С20); · двойная связь в ненасыщенных жирных кислотах располагается между 9 и 10 атомами углерода; · в полиненасыщенных жирных кислотах двойные связи разделены метиленовыми группами (СН=CH-CH2-CH=CH), то есть являются несопряжёнными; · двойные связи находятся в цис-конформации, что приводит к изгибу углеводородной цепи. 9.4.2. Большинство липидов в мембранах млекопитающих представлены фосфолипидами, гликосфинголипидами и холестеролом. Фосфолипиды в составе мембран подразделяются на две группы: глицерофосфолипиды и сфингомиелины. Глицерофосфолипиды – представляют собой сложные эфиры трёхатомного спирта глицерола, двух остатков жирных кислот и фосфорилированного аминоспирта. Общая формула глицерофосфолипида представлена на рисунке 9.5. Наиболее распространённым глицерофосфолипидом мембран является фосфатидилхолин:
В глицерофосфолипидах у второго углеродного атома глицерола обязательно находится остаток ненасыщенной жирной кислоты (в данном случае линолевой).
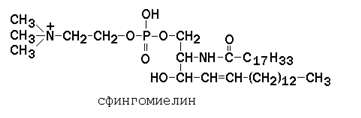
Рисунок 9.5. Общая формула глицерофосфолипидов. Сфингофосфолипиды (сфингомиелины) являются производными аминоспирта сфингозина (рисунок 9.6). Соединение сфингозина и жирной кислоты получило название церамид.
Рисунок 9.6. Структурные формулы сфингозина и его производных. В сфингомиелинах водород гидроксильной группы у первого углеродного атома в церамиде замещён на фосфохолин. Пример сфингомиелина, содержащего остаток олеиновой кислоты:
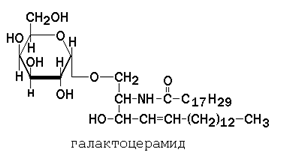
Гликолипиды также являются производными церамида, содержащими один или несколько остатков моносахаридов. Например, цереброзиды содержат в первом положении остаток глюкозы или галактозы:
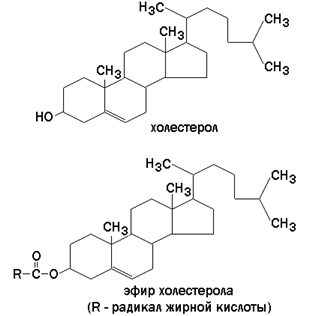
а ганглиозиды содержат цепочку из нескольких остатков сахаров, одним из которых обязательно является сиаловая кислота. Холестерол (рисунок 9.7) – одноатомный циклический спирт. Это один из главных компонентов плазматической мембраны клеток млекопитающих, в меньшем количестве может присутствовать также в митохондриях, мембранах комплекса Гольджи, ядерных мембранах. Особенно много его в нервной ткани.
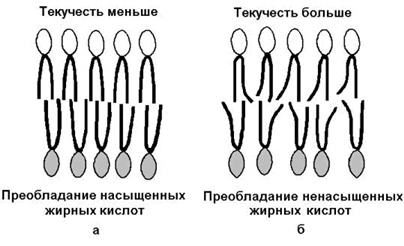
Рисунок 9.7.Структурные формулы холестерола и его эфира. 9.4.3.Как уже было сказано, характерной особенностью мембранных липидов является их амфифильность – наличие в молекуле одновременно гидрофобных и гидрофильных участков. Гидрофобная часть молекулы представлена остатками жирных кислот и боковой цепью сфингозина. Гидрофильные участки представлены в фосфолипидах фосфорилированным спиртом, а в гликолипидах – остатками сахаров. Амфифильность холестерола выражена слабо – циклическая структура и боковой радикал гидрофобны, и только гидроксильная группа гидрофильна. Амфифильность мембранных липидов определяет характер их поведения в водной среде. Слипание гидрофобных участков молекул приводит к образованию упорядоченных замкнутых структур – мицелл, в которых гидрофобные области защищены от воды, а гидрофильные обращены в водную среду. Молекулы холестерола встраиваются между радикалами жирных кислот гидрофобной части бислоя, а его гидроксильная группа примыкает к гидрофильным головкам фосфолипидов. Такая структура, стабилизированная нековалентными гидрофобными взаимодействиями, термодинамически очень устойчива и лежит в основе формирования биологических мембран. 9.4.4.Замкнутый липидный бислой определяет основные свойства мембран: 1) текучесть – зависит от соотношения насыщенных и ненасыщенных жирных кислот в составе мембранных липидов. Гидрофобные цепочки насыщенных жирных кислот ориентированы параллельно друг другу и образуют жёсткую кристаллическую структуру (рисунок 9.8, а). Ненасыщенные жирные кислоты, имеющие изогнутую углеводородную цепь, нарушают компактность упаковки и придают мембране бóльшую жидкостность (рисунок 9.8, б). Холестерол, встраиваясь между жирными кислотами, уплотняет их и повышает жёсткость мембран.
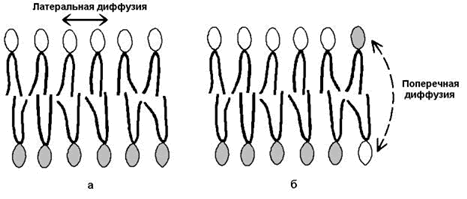
Рисунок 9.8. Влияние жирнокислотного состава фосфолипидов на текучесть липидного бислоя. 2) латеральная диффузия – свободное перемещение молекул относительно друг друга в плоскости мембран (рисунок 9.9,а).
Рисунок 9.9.Виды перемещений фосфолипидных молекул в липидном бислое. 3) ограниченная способность к поперечной диффузии (переходу молекул из наружного слоя во внутренний и наоборот, см. рисунок 9.9, б), что способствует сохранению асимметрии – структурно-функциональных различий наружного и внутреннего слоёв мембраны. 4) непроницаемость замкнутого бислоя для большинства водорастворимых молекул.
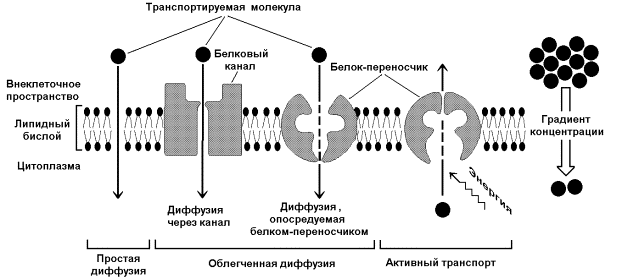
50. Транспорт веществ через мембраны. Механизмы переноса веществ. Транспортные АТФазы. Роль К+,Na+-АТФазы в поддержании трансмембранного потенциала и возбудимости мембраны. Одна из главных функций мембран – участие в переносе веществ. Этот процесс обеспечивается при помощи трёх основных механизмов: простой диффузией, облегчённой диффузией и активным транспортом (рисунок 9.10). Запомните важнейшие особенности этих механизмов и примеры транспортируемых веществ в каждом случае.
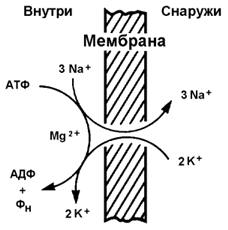
Рисунок 9.10. Механизмы транспорта молекул через мембрану Простая диффузия - перенос веществ через мембрану без участия специальных механизмов. Транспорт происходит по градиенту концентрации без затраты энергии. Путём простой диффузии транспортируются малые биомолекулы – Н2О, СО2, О2, мочевина, гидрофобные низкомолекулярные вещества. Скорость простой диффузии пропорциональна градиенту концентрации. Облегчённая диффузия - перенос веществ через мембрану при помощи белковых каналов или специальных белков-переносчиков. Осуществляется по градиенту концентрации без затраты энергии. Транспортируются моносахариды, аминокислоты, нуклеотиды, глицерол, некоторые ионы. Характерна кинетика насыщения – при определённой (насыщающей) концентрации переносимого вещества в переносе принимают участие все молекулы переносчика и скорость транспорта достигает предельной величины. Активный транспорт – также требует участия специальных белков-переносчиков, но перенос происходит против градиента концентрации и поэтому требует затраты энергии. При помощи этого механизма через клеточную мембрану транспортируются ионы Na+, K+, Ca2+, Mg2+, через митохондриальную – протоны. Для активного транспорта веществ характерна кинетика насыщения. 9.5.2.Примером транспортной системы, осуществляющей активный транспорт ионов, является Na+,K+-аденозинтрифосфатаза (Na+,K+-АТФаза или Na+,K+-насос). Этот белок находится в толще плазматической мембраны и способен катализировать реакцию гидролиза АТФ. Энергия, выделяемая при гидролизе 1 молекулы АТФ, используется для переноса 3 ионов Na+ из клетки во внеклеточное пространство и 2 ионов К+ в обратном направлении (рисунок 9.11). В результате действия Na+,K+-АТФазы создаётся разность концентраций между цитозолем клетки и внеклеточной жидкостью. Поскольку перенос ионов неэквивалентен, то возникает разность электрических потенциалов. Таким образом, возникает электрохимический потенциал, который складывается из энергии разности электрических потенциалов Δφ и энергии разности концентраций веществ ΔС по обе стороны мембраны.
Рисунок 9.11.Схема Na+, K+-насоса. |
||||||||||
|
|
Последнее изменение этой страницы: 2018-05-10; просмотров: 253. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |