|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
Распространены две гипотезы о механизмах полового отбораСогласно гипотезе «хороших генов» самка «рассуждает» следующим образом: «Если этот самец, несмотря на его яркое оперение и длинный хвост, каким-то образом умудрился не погибнуть в лапах хищника и дожить до половой зрелости, то, следовательно, он обладает хорошими генами, которые позволили ему это сделать. Значит, его стоит выбрать в качестве отца для своих детей: он передаст им свои хорошие гены». Выбирая ярких самцов, самки выбирают хорошие гены для своих потомков. Согласно гипотезе «привлекательных сыновей» логика выбора самок несколько иная. Если яркие самцы, по каким бы то ни было причинам, являются привлекательными для самок, то стоит выбирать яркого отца для своих будущих сыновей, потому что его сыновья унаследуют гены яркой окраски и будут привлекательными для самок в следующем поколении. Таким образом, возникает положительная обратная связь, которая приводит к тому, что из поколения в поколение яркость оперения самцов все более и более усиливается. Процесс идет по нарастающей до тех пор, пока не достигнет предела жизнеспособности. В выборе самцов самки не более и не менее логичны, чем во всем остальном их поведении. Когда животное чувствует жажду, оно не рассуждает, что ему следует попить воды, для того чтобы восстановить водно-солевой баланс в организме — оно идет на водопой, потому что чувствует жажду. Точно так же и самки, выбирая ярких самцов, следуют своим инстинктами — им нравятся яркие хвосты. Все те, кому инстинкт подсказывал иное поведение, все они не оставили потомства. Таким образом, мы обсуждали не логику самок, а логику борьбы за существование и естественного отбора — слепого и автоматического процесса, который, действуя постоянно из поколения в поколение, сформировал все то удивительное разнообразие форм, окрасок и инстинктов, которое мы наблюдаем в мире живой природы. 
· Приведите примеры признаков, которые могли возникнуть у живых организмов в результате действия полового отбора. · Заполните сравнительную таблицу
Творческая роль отбора
Изоляция —это нарушение панмиксии и потока генов. Изоляции как фактору, увеличивающему разнообразие организмов, придавали большое значение М. Вагнер, А. Гумбольдт, Ж. Бюффон, П. Л. Мопертюи, Ж. Кювье, А. Р. Уоллес, Ч. Дарвин. "Изоляция также является важным элементом в процессе изменения видов посредством естественного отбора", — писал Дарвин. Он определил изоляцию "как препятствие к скрещиванию". Изоляция есть прекращение потока генов (миграции и скрещивания) географическими преградами, особенностями строения, физиологии, поведения организмов. Поэтому выделяют два типа изоляции — географическую и биологическую. Географическая изоляция — это пространственная, территориальная, климатическая изоляция, возникающая вследствие прекращения миграции (потока генов) и панмиксии географическими преградами. В качестве географических преград могут выступать океанические и морские проливы, реки для сухопутных организмов и суша - для водных. Эффективность океанических и морских проливов известна давно. Так, А. Р. Уоллес обнаружил значительные отличия между островами Бали и Ломбок по биоте: пролив между ними отделяет ориентальную фауну от австралийской. Дарвин изучал результаты пространственной изоляции на Галапагосских островах. Крупные реки часто выступают в качестве географической преграды для сухопутных организмов. Так, Днепр есть граница ареалов двух видов сусликов: на правом берегу обитает крапчатый, а на левом — серый. Для донных морских организмов (бентоса) непреодолимой преградой есть большие океанические преграды и большие океанические хребты. Для абисальных животных непреодолимой преградой выступают неглубокие участки моря. Суша выступает в качестве непреодолимой преграды для рыб и водных беспозвоночных. Панамский перешеек сформировался 2-5 млн. лет назад, когда произошло сближение Северной и Южной Америк. После образования перешейка прежде единые популяции начали дивергировать, и из общей ихтиофауны сформировались атлантическая и тихоокеанская. Из исследованных 1200 видов рыб только 6 % встречаются по обе стороны перешейка, а остальные — отличаются. Водораздел Уральского хребта разграничивает ареалы тритонов: до Уральских гор встречается европейский тритон, за Уралом обитает сибирский тритон. В качестве изолирующего фактора могут выступать климатические преграды. Так, заяц-беляк имеет ареал в лесной зоне, а заяц-русак — в степной. Биологическая изоляция — это биологические барьеры межпопуляционному скрещиванию. Известны два механизма репродуктивной изоляции: презиготические и постзиготические. Презиготические механизмы препятствуют скрещиванию индивидов различных популяций и тем самым исключают возможность появления гибридного потомства. В презиготической изоляции выделяют следующие формы: Экологическая изоляция — изоляция вследствие экологического разобщения. Популяции живут на общей территории, но в различных местах обитания и поэтому друг с другом не встречаются. В горах обычны два вида традесканции: один на скалистых вершинах, другой — в тенистых лесах. Временная изоляция — изоляция вследствие разновременности половой активности или цветения. Максимум кладок серебристой чайки приходится на последнюю треть апреля, а у восточной клуши - не раньше середины мая. Этологическая изоляция — неспаривание вследствие различий в сексуальном поведении (в ухаживании, пении, танцах, свечении, демонстрациях). Брачная окраска, поведение и сигналы самцов воспринимаются только самками того же вида. У млекопитающих важную роль играют химические сигналы. Механическая изоляция — безрезультатность спаривания вследствие разного строения половых органов. Межвидовые спаривания у дрозофилы приводят к травмам и даже к смерти партнеров. Шалфеи различаются строением цветка и поэтому опыляются разными видами пчел. Гаметическая изоляция — отсутствие таксиса между гаметами или же гибель микрогамет в половых путях самки или в рыльцах цветков. Постзиготическая репродуктивная изоляция возникает вследствие: · нежизнеспособность гибридов: зигота развивается в гибрид, обладающий пониженной жизнеспособностью (гибнет зародыш на разных стадиях развития, гибнет молодой организм, гибрид не достигает половой зрелости); · стерильность гибридов: гибриды жизнеспособны, но они не образуют полноценных гамет; · вырождение гибридов — разрушение гибридов: гибриды дают потомков, жизнеспособность и плодовитость которых понижена. У растений репродуктивная изоляция заключается в следующем: · Пыльца другого вида не прорастает на рыльцах цветков другого вида. · Пыльца прорастает, но пыльцевые трубки растут медленно. · Оплодотворение происходит, но зародыш гибнет на разных стадиях эмбриогенеза и жизнеспособное семя не образуется. · Пыльники у гибридов недоразвиты, либо они не открываются. · Происходит нарушение мейоза при образовании гамет. · Составьте схему «Виды изоляции»
· Каково значение изоляции?
Тема 4. Микроэволюция Вид. Критерии вида
Впервые термин “вид” был применен Аристотелем (384-322 д.н.э.) для обозначения общности сходных между собой организмов, отличающихся от других подобных общностей. В сочинениях Аристотеля понятие вид имело больше логический смысл, чем биолого-таксономический. Тем не менее, в его сочинениях было обозначено 500 видов животных. Ученик Аристотеля Теофраст(371-286 д.н.э.) разработал систему растений, аналогичную системе Аристотеля. В своей многотомной сводке “Общая история растений”, издававшейся с 1686 по 1704 гг. англ. биолог Джон Рей впервые ввел понятие вид как категорию, лежащую в основе систематики. Рей предложил называть видом (species) совместно размножающуюся морфологически сходную совокупность особей, которая в потомстве воспроизводит свои морфологические признаки. «Подобное рождает подобное». Он же ввел бинарную номенклатуру, которая впоследствии была утверждена благодаря трудам К.Линнея (1707-1778). Карл Линней впервые сформулировал следующие положения: 1. Вид- универсальная и объективная категория. Т.е. виды реально существуют в природе и нет ни одного организма, который не принадлежал бы к какому-нибудь виду. 2. Вид – основная категория систематики. Основной критерий вида – морфологическая дискретность. Т.е. особи одного вида не должны различаться более, чем различаются дети одних родителей) Виды устойчиво воспроизводят свои признаки в потомках, в то же время четко отграничены друг от друга. Устойчивость формы- основная сущность вида. 3. Поскольку форма устойчива, виды неизменны. Их столько, сколько создал бог. К.Линней создал типологическую концепцию вида. Согласно этой концепции принадлежность особей к тому или другому виду производится путем сравнения её признаков с морфологическим типовым экземпляром - голотипом или с типовой серией – паратипом. Совершенно иной подход развивал Ж. Б. Ламарк, который на основе своей теории эволюции отрицал существование видов – номиналистическая концепция -от слова nomen – имя. Виды – это не реальные единицы, а только названия, придуманные для удобства различения временных морфологически сходных групп. Вопрос о реальности вида вновь возник после трудов Ч. Дарвина. Дарвин признавал реальность вида, но не обсуждал этот вопрос. Его главной целью было – доказательство эволюции видов Он всегда подчеркивал, что обособленность видов появляется в ходе их эволюции. Вид – это резко выраженная разновидность, а разновидность – это начинающийся вид. Это означает, что виды возникают под действием отбора, в процессе дивергенции постепенно обособляются, приобретают дискретность, и тем самым – реальность. Своими трудами Ч.Дарвин доказал изменяемость видов и их историчность. В связи с этим возникла новая проблема – вопрос о сущности, границах и реальности вида. Этот вопрос в ХIХ веке это представлял собой значительную трудность. При его разрешении здесь наметились два направления: 1.Вид - элементарная и далее неподразделимая единица. Поскольку большинство видов можно было разложить на разновидности и расы, такие виды нельзя был принимать как далее не подразделимые элементарные единицы. Поэтому эти систематики выделяли в качестве видов группы особей, сохраняющие в потомстве свои признаки. Основателем такого типологического подхода является ботаник А. Жордан. Вид это элементарная неизменная и не эволюционирующая группа особей с четкими и дискретными морфологическими различиями, предаваемыми по поколениям. Впоследствии такие понятия Г.де Фриз назвал – жорданонами(euspecies). Такой подход на практике систематиков привел к видодробительству. Пример: А.Жордан у растения крупка весенняя выделил 300 жорданонов, возводимых им на ранг вида. Это направление получило подтверждение в исследованиях генетиков в виде существования чистых мутантных линий. Оно защищало свои позиции, основываясь на исследованиях ранних генетиков. 2. Вид как сложная иерархическая система. На основе морфологического сходства близких форм эти систематики стремились, наоборот, к видоукрупнительству. Впоследствии Г.де - Фриз назвал их линнеонами(conspecies), или линнеевскими видами. Это направление не стояло на месте и постепенно видоизменилось в другое направление, называемое эколого-географическим. Оно основывалась на биогеографических критериях и также ввело новый критерий вида – ареалогический. Видом считать совокупность особей, распространенных в пределах обширного ареала, сходных между собой и в течение продолжительного времени не меняющих свой морфологический облик. Со временем это привело к развитию концепции политипического морфологического вида, основателем которого является А.П.Семенов-Тян-Шанский (1910), который впервые ввел иерархию видовых категорий: вид(species)-подвид(subspecies)-племя(natio)-морфа(morfa)-аберрация aberratio). Сформировалось представление о линневском виде как иерархической системы пространственно или сезонно различающихся групп. Решение проблемы реальности политипического вида было достигнуто Н.И. Вавиловым в его труде «Линнеевский вид как система». В этой работе были даны материалы по изучению гомологических рядов изменчивости и центрам происхождения культурных растений. На основе синтеза этих данных Н. И. Вавилов показал, что вид является сложную иерархическую систему, состоящую из множества взаимоподчиненных элементов, различающихся морфологически, генетически и экологически, формирующуюся в ходе эволюции и обладющую единым ареалом. Н.И.Вавилов впервые показал, что внутривидовое разнообразие является важным условием существования вида и для его эволюции. То есть эволюционная динамика вида неразрывно связана с динамикой его существования, что делает вид относительно стабильным и реально существующим. В 1940-х гг была создана СТЭ, утвердившая понятия микроэволюция и её элементарные факторы, в числе которых большое значение имеет изоляция и различные формы естественного отбора. Было внесено представление о популяционной структуре вида и его пространственной структуре. Таким образом, была сформирована новая биологическая концепция вида. Перечень научных монографий, иллюстрирующий ход становления данной концепции таков: Ф.Добржанский «Генетика и происхождение видов»,1937; Э.Майр «Систематика и происхождение видов», 1942; Э.Майр, Линсли, Юзингер «Принципы и методы современной систематики», 1953; Э.Майр. «Зоологический вид и эволюция», 1968. Согласно новой концепции виды состоят не из отдельных независимых особей, а из популяций, а основным критерием вида стала его генетическая обособленность, в становлении которых играет большую роль изоляция. При этом решающим условием различения видов является не плодовитость при скрещивании, а репродуктивная изоляция, которая может принимать различные значения и формы. Биологическая концепция вида считает вид как репродуктивное сообщество, комплекс популяций обменивающихся между собой генами и не обменивающиеся ими с другими подобными сообществами. Иными словами, видовая принадлежность особи, или группы особей(популяции) определяется не по степени сходства с типом данного вида, а по наличию любых признаков, подразумевающий обособленность генофонда вида. Вид - это генетически изолированная группа особей. Биологическая концепция теоретически обосновала новый основной критерий вида – генетический. Вид оказался сложной генетической системой. В 1970-80-х гг. биологическая концепция прошла очередной этап развития, что привело к формированию признанной в настоящее время современной концепции политипического вида. В настоящее время всеми в общих чертах признается, что вид представляет собой качественный этап процесса эволюции. Вид – это генетически устойчивая система скрещивающихся популяций, (обладающих общей эволюционной судьбой и формирующих общий ареал), особи которых объединены возможностью скрещивания друг с другом, обладают сходством морфофизиологических признаков и экологическим своеобразием, занимая в экосистемах совокупности экологических ниш. Согласно современным представлениям вид – это главный этап эволюции. Как главный этап эволюции он имеет определенные свойства- с одной стороны – это результат эволюции, а с другой это этап эволюционного процесса. 1. Как результат эволюционного процесса вид целостная, приспособленная к данной среде, генетически обособленная от всех других таких же видов, устойчивая система. 2. Как этап эволюционного процесса вид динамичен, изменчив, имеет расплывчатые границы. Вид состоит из множества популяций, каждая из которых является совершенно совершенно самостоятельной единицей с собственной эволюционной судьбой. Однако генофонды популяций обмениваются генетическим материалом, между ними существует поток генов, который служит основой для формирования единой сложной иерархической системы популяций. Это создает генофонд вида - наименьшую далее неделимую устойчивую генетическую систему. Структура вида Любой вид распадается на относительно изолированные группы особей, объединенных тесным генетическим родством (демы- у животных, биотипы – у растений). У животных такие группы демов могут объединять микропопуляции, а последние объединяются в популяции. Одиночные популяции или их группы могут образовывать расы и подвиды. У растений группы демов могут образовывать популяции, а последние могут образовавать экотипы и подвиды. Вообще, при описании внутривидовой структуры не только в ботанике и зоологии, на и в различных биологических дисциплинах используются разные понятия. Единого мнения нет даже внутри одной дисциплины, даже в зоологии и ботанике. Однако всеми признается, что любой вил - сложная система с многоуровневой иерархией биохорологических групп. Большинство видов политипично, т.е. состоят из ряда иерархически соподчиненных внутривидовых групп. Например, если у животных виды состоят из подвидов, популяций и микропопуляций, то у растений это может выглядеть сложнее. Так различаются понятия: · Полувид –географическая или экологическая раса, почти достигшая состояния молодого вида. · Подвид – сформированная экологическая или географическая раса. · Местная популяция – относительно обособленное поселение, устойчиво занимающее определенную территорию и способное к воспроизведению · Экоэлемент – внутрипопуляционная форма, связанная со спаянным нерасщепляющимся генотипом. Может выходить из состава популяции и существовать как самостоятельный экотип. · Морфобиологическая группа – группа организмов популяции, имеющая одинаковую или различную наследственную основу, различимая морфологически, имеющая сходный путь развития и одинаково реагирующая на условия среды. · Биотип – элементарная единица генетической структуры популяции – группа организмов, имеющих почти тождественный генотип и отличающаяся от других таких же групп хотя бы одной мутацией. Вид - исторически сложившаяся совокупность особей, занимающих определенный ареал, сходных по морфофизиологическим свойствам, способных свободно скрещиваться между собой и давать плодовитое потомство.
Морфы —популяции или субпопуляции одного вида, которые отличаются друг от друга среди прочего фенотипами.Например, субпопуляции форели Salmo trutta морфа trutta и Salmo trutta морфа fario. Эти две субпопуляции принадлежат к одному и тому же виду и могут скрещиваться. Еще одним примером может служить арктический голец (Salvelinus alpinus), которые в одном и том же озере развились в разные морфы, питающиеся планктоном и рыбой. Питающиеся рыбой морфы питаются всеми обитающими там рыбами в среднем и верхнем слое воды, в том числе поедающими планктон морфами. Напротив, питающиеся планктоном морфы питаются зоопланктоном в верхнем слое воды или другими беспозвоночными. Они меньше размером, растут медленнее и получают меньше потомство.Разделение популяций, особенно изолированных друг от друга, на морфы можно рассматривать в некоторых случаях в качестве предшественника симпатричного видообразования.
· У большинства млекопитающих и многих видов птиц самцы более крупные и тяжёлые, чем самки. У земноводных и членистоногих самки, как правило, крупнее самцов. У черепах и у змей самки крупнее самцов; у ящериц, напротив, как правило, самцы крупнее самок; исключение составляют гекконы, у которых большая величина также свойственна самкам. · Борода у мужчин, грива у львов или бабуинов. · Цвет оперения у птиц, особенно у утиных. · Характерные наросты или дополнительные образования, такие как рога у оленевых, гребешок у петухов. · Бивни у самцов индийского слона, более крупные клыки у самцов моржей и кабанов. · Некоторые животные, прежде всего рыбы, демонстрируют половой диморфизм только во время спаривания. Согласно одной из теорий, половой диморфизм выражен тем больше, чем различнее являются вклады обоих полов в уход за потомством. Также он является показателем уровня полигамии. · Как отличить представителей разных видов? · Какие критерии для этого можно использовать? Заполните таблицу. Критерии вида
Видообразование · Что такое видообразование? · Одинакова ли скорость видообразования у различных видов живых организмов? Почему? · От чего зависит скорость видообразования? · Изучите материал и заполните сравнительную таблицу.
Видообразование в типичных случаях заключается в разделении первоначально единого вида на два или более новых. Это связано с возникновением межпопуляционных изоляционных барьеров и углублением различий между генофондами популяций под действием естественного отбора вплоть до генетической изоляции. Такой процесс, ведущий к увеличению количества видов, называют дивергентным или истинным видообразованием.
Образование видов в результате скрещивания популяций разных видов на территории пересечения их ареалов называется гибридогенным видообразованием.По некоторым данным, гибридогенное происхождение имеют не менее трети всех видов цветковых растений. Примером гибридогенного вида является рябинокизильник, распространенный в лесах центральной Сибири. Кроме того, межвидовые гибриды растений могут размножаться вегетативным путем и распространяться за счет этого.Для животных полиплоидия не столь характерна, поэтому гибридогенное видообразование встречается реже, однако известны виды кавказских скальных ящериц гибридогенного происхождения, а также амфибий и рыб. Некоторые виды кавказских ящериц, имеющих гибридогенное происхождение, триплоидны и размножаются с помощью партеногенеза. Выделяют 2 разновидности дивергентного видообразование: аллопатрическое и симпатрическое. При аллопатрическом видообразовании, называемом также географическим, препятствия к скрещиванию первично обусловлены пространственным разобщением популяций. Генетическая изоляция развивается вторично. Аллопатрическое видообразование происходит медленно и дает виды, как правило, отличающиеся по морфофизиологическому критерию от вида-родоначальника. Характерно в основном для животных. Например, в Австралии существовал один вид попугайчиков рода Pachycephala. В условиях засушливого периода единый ареал разделился на западную и восточную зоны. Со временем особи При симпатрическом видообразовании новый вид образуется внутри ареала исходного вида. С самого начала изоляция является генетической. Такое положение создается в результате полиплоидии вследствие нарушений нормального хода мейоза, при крупных хромосомных перестройках или межвидовой гибридизации. Симпатрический путь относительно быстрый и дает виды, близкие к исходному по морфофизиологическим показателям.Симпатрическое видообразование на основе полиплоидии характерно для растений. Например, разные виды пшениц составляют ряд с наборами 14, 28, 42 хромосомы. В клетках дикого хлопчатника 26 хромосом, культурного — 52. Культурная слива возникла путем гибридизации терна с алычой. Симпатрический путь видообразования у паразитов часто связан с освоением популяцией новых хозяев. Анализ генного состава и межхромосомных различий между человеком и человекообразными обезьянами дает повод предположить, что разделение этих двух ветвей могло идти симпатрическим путем.Классическим примером симпатрического видообразования является сверхбыстрая дивергенция рыб-цихлид больших африканских озёр — Малави, Танганьика и Виктория. Озёра имеют разный возраст, но все они относительно молоды. Каждое озеро изначально было заселено небольшим числом видов рыб, которые стали быстро дивергировать, причём эволюция в каждом озере шла в значительной мере независимо. В озере Танганьика 250 видов цихлид образовалось за 12−15 млн лет, в Малави — 500 видов менее чем за 5 млн лет. Рекордная скорость видообразования зафиксирована в самом молодом озере, Виктории, где на формирование 500 эндемичных видов ушло, по разным оценкам, от 15 до 100 тысяч лет. Как обычно бывает в подобных случаях, освоение сходных ниш вело к независимому появлению сходных жизненных форм в разных озёрах.
|
||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2018-05-10; просмотров: 476. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |
 Систематика- наука о многообразии живых организмов и распределении их по группам на основании (эволюционного) родства.
Систематика- наука о многообразии живых организмов и распределении их по группам на основании (эволюционного) родства.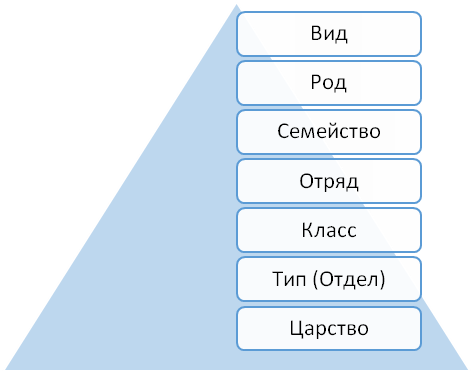
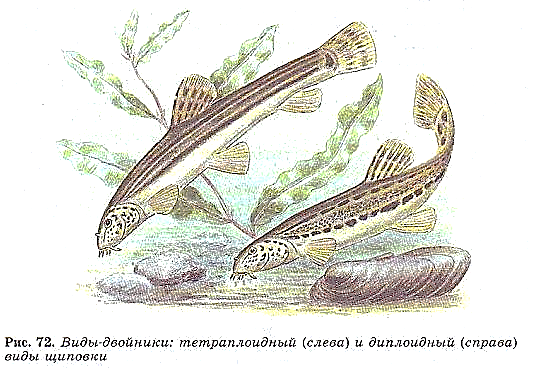
 Виды-двойники(Майр, 1942) - морфологически сходные или идентичные, репродуктивно изолированные природные популяции.Видов-двойников очень много среди насекомых, моллюсков, ракообразных и простейших. Меньше их среди позвоночных, хотя виды-двойники открыты у землероек и грызунов, рыб и лягушек. Классический случай — комплекс малярийных комаров в Европе. Считалось, что возбудителя малярии переносит единственный вид комара — Anopheles maculipennis. Но изучение распространения комаров показало, что они обычны и в тех районах Европы, где малярии нет. Кроме того, были установлены различия в питании комаров: в одних районах они питаются на домашних животных, в других же — на человеке. В одних случаях личинки комаров развиваются только в пресной воде, в других — в солоноватой.Виды-двойники есть среди муравьев, жуков-листоедов и короедов, сверчков, кобылок и других насекомых. В том числе этим и объясняется пересмотр и уточнение систематического положения многих видов, приводящих в одних случаях к упрощению систематики ряда групп, а в других — к увеличению числа описываемых видов.
Виды-двойники(Майр, 1942) - морфологически сходные или идентичные, репродуктивно изолированные природные популяции.Видов-двойников очень много среди насекомых, моллюсков, ракообразных и простейших. Меньше их среди позвоночных, хотя виды-двойники открыты у землероек и грызунов, рыб и лягушек. Классический случай — комплекс малярийных комаров в Европе. Считалось, что возбудителя малярии переносит единственный вид комара — Anopheles maculipennis. Но изучение распространения комаров показало, что они обычны и в тех районах Европы, где малярии нет. Кроме того, были установлены различия в питании комаров: в одних районах они питаются на домашних животных, в других же — на человеке. В одних случаях личинки комаров развиваются только в пресной воде, в других — в солоноватой.Виды-двойники есть среди муравьев, жуков-листоедов и короедов, сверчков, кобылок и других насекомых. В том числе этим и объясняется пересмотр и уточнение систематического положения многих видов, приводящих в одних случаях к упрощению систематики ряда групп, а в других — к увеличению числа описываемых видов.

 Половой диморфизм— анатомические различия между самцами и самками одного и того же биологического вида, исключая различия в строении половых органов.
Половой диморфизм— анатомические различия между самцами и самками одного и того же биологического вида, исключая различия в строении половых органов.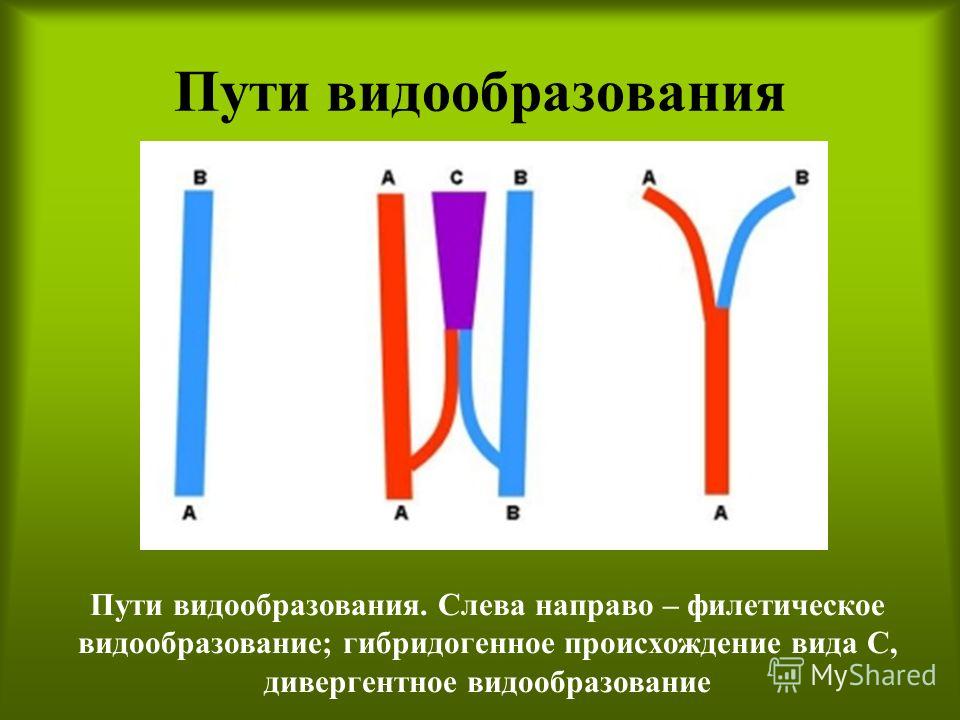 Видообразование – это процесс образования видов осуществляется в результате взаимодействия элементарных эволюционных факторов.
Видообразование – это процесс образования видов осуществляется в результате взаимодействия элементарных эволюционных факторов.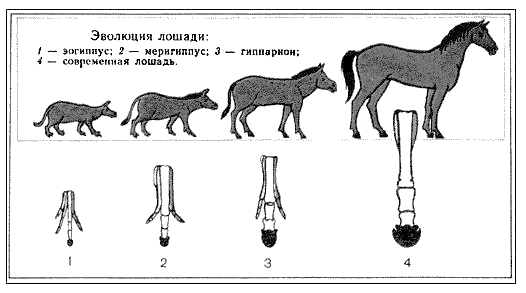 Выделяют также филетическое видообразование. Оно заключается в постепенном превращении во времени одного вида в другой. Этот способ наблюдается, если изменения условий захватывают весь ареал. Вид изменяется в череде поколений (например, ряд ископаемых европейских слонов, лошадей), превращается в новый вид. Границы между отдельными видами в филетическом ряду форм провести невозможно – она всегда будет условной. В «чистом виде» филетическая эволюция, видимо, возможна лишь как идеализированное и упрощенное отражение эволюционного процесса на одномизотрезков жизни вида (фратрии).
Выделяют также филетическое видообразование. Оно заключается в постепенном превращении во времени одного вида в другой. Этот способ наблюдается, если изменения условий захватывают весь ареал. Вид изменяется в череде поколений (например, ряд ископаемых европейских слонов, лошадей), превращается в новый вид. Границы между отдельными видами в филетическом ряду форм провести невозможно – она всегда будет условной. В «чистом виде» филетическая эволюция, видимо, возможна лишь как идеализированное и упрощенное отражение эволюционного процесса на одномизотрезков жизни вида (фратрии).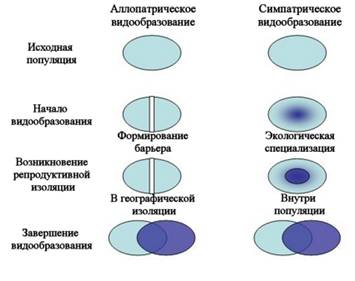 двух популяций приобрели морфофизиологические различия, которые сделали невозможным скрещивание, когда ареал вновь стал общим. Произошло образование из одного предкового вида двух новых.
двух популяций приобрели морфофизиологические различия, которые сделали невозможным скрещивание, когда ареал вновь стал общим. Произошло образование из одного предкового вида двух новых.