Значение активных сил в формировании мембранного потенциала.
Основы электрофизиологии сердца
Основными функциями клеток сердца являются: функция автоматизма, функция возбудимости, функция рефрактерности, функция проводимости, и функция сократимости.
· Автоматизм – это способность клеток автоматически вырабатывать электрические импульсы.
· Возбудимость - способность клеток возбуждаться электрическими импульсами.
·Рефрактерность - временная невозбудимость клеток, когда электрические импульсы неспособны вызвать возбуждение.
· Проводимость - способность клеток проводить электрические импульсы.
· Сократимость - способность клеток сокращаться при возбуждении.
Функция автоматизма.
Повторю, автоматизм – способность клеток автоматически вырабатывать электрические импульсы.
Клетки, автоматически вырабатывающие электрические импульсы, образуют центры автоматизма.
Выделяют центры автоматизма I, II, III порядка.
Центром автоматизма I порядка являются Р-клетки синусового узла, который находится в верхнезадней части правого предсердия - между устьями полых вен.
В нем в норме возникают импульсы частотой у взрослых - 60-80 в 1 минуту и до 140 у маленьких детей.
Центром автоматизма II порядка являются Р-клетки правого и левого предсердия, Р-клетки АВ-соединения (в NH-зоне). Частота импульсов ц центрах II порядка меньшая, обычно 40-60 в 1 минуту.
Центрами автоматизма III порядка являются клетки пучка Гиса и волокон Пуркинье. Частота импульсов в них - 20-40 в 1 минуту.
В норме водитель ритма – синусовый узел (номотопный центр автоматизма)\
Другие центры автоматизма являются ненормальными - гетеротопными или эктопическими.
Очень важно запомнить, что в норме автоматизм эктопических центров подавляется синусовым узлом и нижележайшие центры являются запасными, страхующими сердце от катастрофы. Автоматизм эктопических центров проявляется при угнетении автоматизма синусового узла или при повышении автоматизма эктопических центров - при нарушении проведения электрических импульсов от синусового узла по типу блокады, при нарушении проведения электрических импульсов от синусового по типу re-entrу и др. Обычные клетки миокарда желудочков не обладают способностьч автоматизма. Надо также знать, что синусовый узел и АВ-узел имеют симпатическую и парасимпатическую иннервацию и, следовательно подвержен всевозможным экстракардиальным влияниям.
Функция возбудимости.
Возбудимость - это способность сердца возбуждаться под влиянием импульсов. Функцией возбудимости обладают клетки как проводящей системы сердца, так и сократительного миокарда. Возбуждение сердечной мышцы сопровождается возникновением трансмембранного потенциала действия (ТМПД) и в конечном счете - электрического тока.
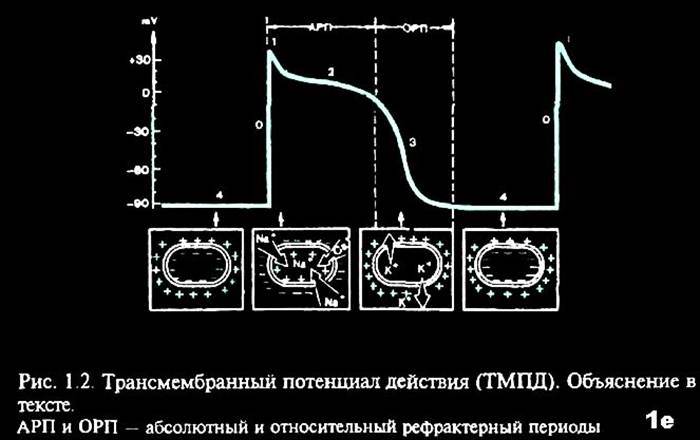
В разные фазы ТМПД возбудимость мышечного волокна при поступлении нового импульса различна. В начале ТМПД клетки полностью невозбудимы, или рефрактерны к дополнительному электрическому импульсу (1,2). Это так называемый абсолютный рефрактерный период миокардиального волокна, когда клетка вообще неспособна отвечать новой активацией на какой-либо дополнительный электрический стимул В конце ТМПД имеет место относительный рефрактерный период, во время которого нанесение очень сильного дополнительного стимула может привести к возникновению нового повторного возбуждения клетки, тогда как слабый импульс остается без ответа (3). Во время диастолы полностью восстанавливается возбудимость миокардиального волокна, а его рефрактерность отсутствует (4).
Значение активных сил в формировании мембранного потенциала.
Перемещение ионов происходит путем диффузии. Активный транспорт осуществляется за счет Nа+-К+ насоса (Р. Дин - 1941 г.). Nа+-К+ насос осуществляет движение ионов против градиента концентрации (К+ внутрь, Nа+ - наружу). Для работы насоса требуется энергия, которая образуется при расщеплении АТФ под влиянием АТФ-азы, которая активизируется при изменении концентрации К+ и Nа+, что происходит постоянно, поэтому Nа+-К+ насос работает постоянно. Согласно Дину движение ионов осуществляется за счет молекул переносчиков (белки внутри клеточных мембран). После выполнения функции Х-белок (переносчик ионов К+), благодаря энергии АТФ, меняет свою структуру и превращается в У-белок (переносчик ионов Nа+). Nа+-К+ насоса неодинакова при различных состояниях. В состоянии покоя на 3 иона Nа+ приходится 2 иона К+. При изменении состояния клетки меняется активность Nа+-К+ насоса.

| Итак, в состоянии покоя за счет выхода ионов К+ из клетки наружная поверхность клетки заряжена положительно, а внутренняя - отрицательно (по отношению к наружной поверхности). Это состояние называется поляризация; мембранный потенциал является равновесным калиевым потенциалом; в возникновении мембранного потенциала участвуют другие ионы и активные силы.
Механизм формирования потенциала действия.
Потенциал действия возникает в ткани под влиянием порогового и сверхпорогового раздражителей и является импульсивным возбуждением. Потенциал действия можно так же, как и мембранный потенциал, зарегистрировать трансмембранным способом. Под влиянием пороговых раздражителей изменяется проницаемость клеточной мембраны - повышается для всех потенциалобразующих ионов, но больше всего для ионов Nа+ (в 500 раз). Ионы натрия перемещаются внутрь клетки. Движение ионов натрия внутрь клетки превышает выход ионов К+ из клетки. В результате происходит изменение заряда клеточной мембраны на противоположный, затем происходит постепенное восстановление исходного заряда мембраны.

Компоненты потенциала действия и механизм их возникновения.
При трансмембранном способе регистрации возникает потенциал действия, состоящий из 3-х основных компонентов:
1 компонент: местный (локальный ответ);
2 компонент: пик (спайк);
3 компонент: следовые потенциалы (отрицательный и положительный).
Спайк (пик) - самая постоянная часть. Он состоит из восходящего колена (фаза деполяризации) и нисходящего колена (реполяризация). Остальные компоненты изменчивы и могут отсутствовать.
Местный (локальный) ответ возникает и продолжается до тех пор, пока раздражитель не достигнет пороговой величины. Если раздражитель (его сила) меньше 50-75 % пороговой величины проницаемость мембраны изменяется незначительно и равновесно для всех ионов (неспецифично). После достижения силы раздражителя 50-75 % начинает преобладать натриевая проницаемость, т. к. натриевые каналы освобождаются от ионов Са2+. Происходит снижение мембранного потенциала при достижении пороговой величины разность потенциалов достигает критического уровня деполяризации.
Критический уровень деполяризации (Ек) - это та разность потенциалов, которая должна быть достигнута, чтобы местные изменения перешли в пик потенциала действия. Ек - пороговая величина, при которой местные изменения переходят в распространенные. Ек величина практически постоянная и равна - 40 - -50 мВ. Разность между мембранным потенциалом и пороговой величиной характеризует порог раздражения и отражает возбудимость ткани.
Пик потенциала действиясостоит из следующих фаз.
Фаза деполяризации возникает в результате лавинообразного движения Nа+ внутрь клетки. Этому способствуют две причины: открываются потенциалзависимыеNа+-каналы. В этом случае происходит деполяризация по типу процесса с положительной обратной связью (самоподкрепляющийся процесс).
Освобождение натриевых каналов от Са2+.
Заряд клеточной мембраны сначала снижается до 0 (это собственно деполяризация), а затем меняется на противоположный (инверсия или овершут). Для характеристики фазы деполяризации вводится понятие реверсии - это та разность потенциалов, на которую потенциал действия превышает потенциал покоя.
Р=(потенциал действия) - (мембранный потенциал) 20-30 = 50-60 мВ.
Р (реверсия) - это то количество мВ на которое произошла перезарядка мембраны. Фаза деполяризации продолжается до достижения электрохимического равновесия по Nа+. Затем наступает следующая фаза. Амплитуда потенциала действия не зависит от силы раздражителя. Она зависит от концентрации Nа+ (как снаружи, так и внутри клетки), от количества натриевых каналов, особенностей натриевой проницаемости.
Фаза реполяризациихарактеризуется:
снижением проницаемости клеточной мембраны для Nа+ (Nа-инактивация). Натрий накапливается на наружной поверхности клеточной мембраны;
возрастанием проницаемости мембраны для К+, в результате повышается выход К+ из клетки с увеличением положительного заряда на мембране;
изменением активности Nа+-К+ насоса.
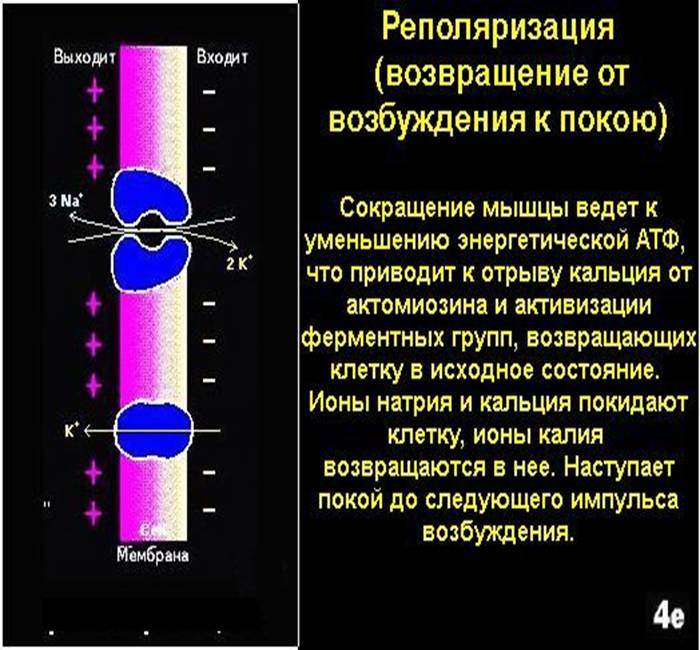
Реполяризация - это процесс восстановления заряда мембраны. Но полного восстановления нет, т. к. возникают следовые потенциалы.
Следовые потенциалы делятся на:
Отрицательный следовой потенциал - замедление реполяризации клеточной мембраны. Это результат проникновения внутрь клетки какого-то количества Nа+, таким образом, отрицательный следовой потенциал - это следовая деполяризация.
Положительный следовой потенциал - увеличение разности потенциалов. Это результат повышенного выхода ионов К+ из клетки. Положительный следовой потенциал - это следовая гиперполяризация. Как только калиевая проницаемость возвращается к исходному уровню - регистрируется мембранный потенциал.
Функция проводимости
Проводимость - способность клеток проводить электрические импульсы
электрические импульсы проводят клетки проводящей системы сердца и кардиомиоциты.
|
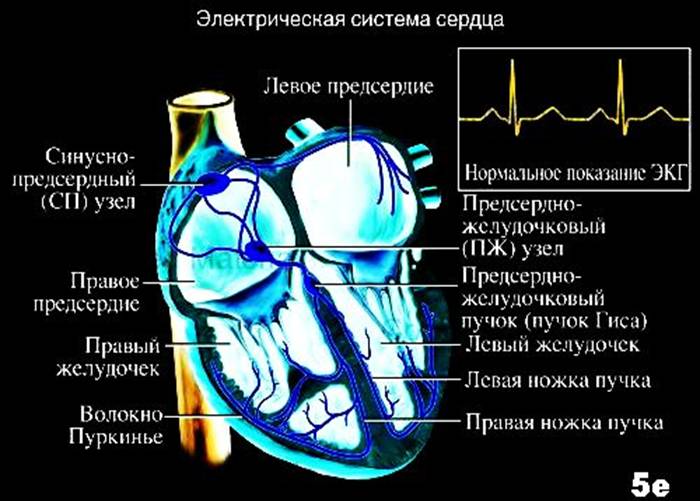
В норме проводящая система сердца для проведения электрических импульсов от синусового узла включает кардиомиоциты предсердий, АВ-узел, пучок Гиса, правая и левая ножки пучка Гиса, волокна Пуркинье.
Скорость проведения импульсов в предсердиях 1м/с, АВ-узле 0,2м/с, пучке Гиса 1м/с, в ножках и волокнах Пуркинье 3-4м/с.
В норме такая проводящая система определяет последовательность проведения возбуждения в сердце синусового узла. От синусового узла электрические импульсы проводятся на кардиомиоциты предсердий.
В предсердиях электрические импульсы проводятся от правого предсердия на левое предсердие по пучку Бахмана, причем все предсердия возбуждаются за 0,1с.
По кардиомиоцитам предсердий электрические импульсы проводятся на АВ-узел.
По АВ-узлу электрические импульсы проводятся с низкой скоростью – происходит задержка проведения. Эта задержка физиологична – в итоге, систола желудочков наступает после систолы предсердий.
От АВ-узла электрические импульсы проводятся на пучок Гиса, ножки пучка Гиса, волокна Пуркинье и далее на кардиомиоциты желудочков.
В желудочках электрические импульсы распространяются от средней части межжелудочковой перегородки на верхушку правого желудочка, затем на верхушку левого желудочка, затем на базальную часть желудочков и перегородки
Все желудочки возбуждаются за 0,1с, причем распространяется от эндокарда к эпикарду.
|
