|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
АРСЕНАЛ ДОЛГОВРЕМЕННОЙ ПАМЯТИ
Как было сказано выше, на пути основного информационного потока в каждой доле таламуса располагаются установочные линзы (рис. 2.5).
Информационные матрицы до линз и после них по биохимической и энергетической структуре близки. На установочных линзах происходит формирование их геометрии в виде спирали, закрученной (по ходу движения) по часовой стрелке. Цепи, проходя (каждая в отдельности) доли таламуса, пополняются биохимической основой и формируются в более плотную спираль. При движении по долям таламуса они вступают во взаимосвязь с центрами вегетативной системы и ядрами таламуса. 
При этом в нижних этажах таламуса цепи делают второй поворот, входя в структуру гипоталамуса. В нижних слоях происходит также считывание значимой информации. Она уходит при помощи субчастиц через ликвор в виде блуждающих импульсов. При схождении двух спиралевидных цепей идет их «опрессовка» и скручивание с получением единой полинуклеотидной матрицы, несущей информацию дневного накопления. Если две разделённые спирали были закручены по часовой стрелке, то Этот процесс можно сравнить с соединением двух половин разорванного листа. На начальной стадии соединения витки спиралей достаточно рыхло прилегают друг к другу, но в дальнейшем они становятся всё более плотными. Поэтому после завершения процесса формирования полинуклеотидная матрица имеет форму конической спирали (улитки), чей острый конец направлен в сторону гипофиза. Хотя информационная доукомплектация уже прошла на 97%, в гипоталамусе набираются последние 3%, которые складываются из следующих поступлений: Процесс доукомплектации и более плотного скручивания витков можно назвать механизмом «опрессовки». Он может длиться до трёх суток. Весь описанный процесс и создаёт дневной информационный комплекс, принимающий овеществлённую форму в виде полинуклеотидной матрицы. Случается, что матрица ещё не реализовалась, но формирующаяся следующая начинает «торопить». В этом случае либо увеличивается скорость обработки первой матрицы, либо она полностью или частично (в зависимости от уровня значимости) разрушается. СТАБИЛИЗИРУЮЩИЕ ОСИ
В первой главе сообщалось, что стабилизирующие оси проходят в больших полушариях (по три в каждом), не пересекая друг друга. Они сходятся в гипоталамусе, подходя к шести его ядрам. В этой области на осях сказывается влияние как программ арсенала памяти, так и ядер гипоталамуса. В процессе заполнения программ арсенала памяти стабилизирующие оси используются незначительно. По своим функциям они являются организующей формацией, диктующей порядок распределения информации. Хотя стабилизирующие оси и не регулируют сам процесс умственной деятельности, но при травмах и других нарушениях головного мозга может происходить их индивидуальное замыкание с отключением энергоснабжения обширных отделов мозга. При этом в дальнейшем происходят и морфологические изменения. Искусственное «отключение» какой-либо из осей чревато последствиями. Для наглядности составим «карту», привязанную к морфологическим структурам мозга. Расположим циферблат часов в плоскости так, чтобы цифра 12 приходилась на район темени, «6» – на район гипофиза, «3» – на мозжечок, а «9» – на лобные доли. Пронумеруем оси справа налево (рис. 2.7, 2.8а, б). Как было сказано ранее, все шесть стабилизирующих осей морфологически однотипны, но различаются своей несущей частотой и контролируемым участком коры. Оси имеют двойственную – вещественно-полевую – природу. Вещественная их часть выглядит как цепь нейронов, собственно же оси образованы, предположительно, белково-полисахаридными молекулами. Стабилизирующие оси – своеобразные «колебательные контуры» мозга. Они осуществляют связь с огромным количеством арсенальных программ, расположенных в корковом и подкорковом веществах мозга.
Во-вторых, оси стабилизируют поступающую дневную информацию. Своей максимально стабильной энергетической структурой оси противостоят информационному хаосу. Он неминуемо возник бы в головном мозге, если бы все программы, «заинтересованные» в получении информации, начали одновременно ею заполняться. Путём специфической энергетической кодировки проходящей через них информации как во время сна, так и в фазу бодрствования, оси управляют количеством одновременно заполняемых информацией программ. Механизм, позволяющий снабжать программы текущей информацией в порядке их максимальной заинтересованности, реализуется с помощью блуждающих импульсов до востребования.
БЛУЖДАЮЩИЕ ИМПУЛЬСЫ
Программы, расположенные в подкорке, и наиболее значимая в них информация своими активными радикалами образуют формирование, похожее на густую сеть. Энергетические импульсы, курсирующие в головном мозге, могут, обогащаясь встречающейся на их пути информацией могут стать основой новой для новой программы . Такие импульсы могут быть запущены сознательно или неосознанно. Курсируя по радикалам арсенальных программ, они могут сопоставлять даже не сопоставимую информацию. Назовем их блуждающими импульсами. Существуют два вида блуждающих импульсов: сторожевые и до востребования. Они являются не только сборщиками информации, но и своеобразными энергетическими аккумуляторами. Блуждающие импульсы автономно могут существовать до одних суток, получая энергию из гипоталамической области. Рассмотрим первый вид блуждающих импульсов – сторожевые. Сторожевые импульсы создаются, как правило, неосознанно. Молекулярная масса этих постоянно циркулирующих по коре больших полушарий частиц невелика. Сторожевые импульсы довольно быстро теряют энергию, поэтому вынуждены пополнять её за счет транспортных соединений, переносимых кровью. Периодически, по мере обесточивания, они разрушаются и создаются снова из транспортных биохимических частиц, которые приносятся с биологическими жидкостями мозга. Сторожевые блуждающие импульсы всегда имеют материальную основу. Обмен информацией в них осуществляется двумя способами. Кора полушарий имеет борозды, где серое вещество очень близко подходит к стабилизирующим осям (до 1 см). В таких местах блуждающие импульсы могут спускаться в борозды и взаимодействовать с осями, воспринимая с них как отдельные информационные кванты, так и целые комплексы. Затем самостоятельно или при подключении других транспортировщиков эта информация может передаваться на арсенальные программы (если считывается максимально значимая информация, что бывает нечасто). «Вытягивание» блуждающими импульсами информации происходит при их гомологичности информационному фрагменту. Импульс имеет до шести специфических окончаний, которые родственны энергетическим нишам фрагментов максимально значимой информации. Таких блуждающих импульсов немного, до 2–3% от всего объёма. Имеется и другой энергетический механизм «перекачивания» информации на импульсы. При совпадении кода импульса с арсенальной программы фрагментарно или полностью дублируется энергоинформационная составляющая и переводится на импульс. Запись информации выражается в изменении углов поворота атомов в материальной структуре блуждающего импульса. Повторяемость опознающих кодовых сочетаний среди блуждающих импульсов незначительна и составляет 10–15%. Сторожевые импульсы имеют две основные функции: энергетической подпитки программ и дополнительного переноса информации с одной программы на другую (с последующим формированием новых программ и их проверкой). Энергетическая матрица новой сформированной программы переносится на стабилизирующие оси. Если квант одного информационного фрагмента плюс сотни тысяч других радикалов сформировали совместно матрицу и она оказалась целесообразной – новая программа будет устойчивой. В противном случае образованная матрица разрушается. Сторожевые импульсы, перемещаясь по программам и собирая с них энергетические слепки, готовят как бы «репродуктивный слепок», который в последующем попадает на одну из шести стабилизирующих осей. На оси происходит его стыковка с энергослепками других сторожевых импульсов, что приводит к формированию матрицы будущей программы в виде энергетического кольца. Если нет дефицита строительных биохимических материалов, а он наблюдается редко, вдоль стабилизирующей оси формируется новая энергоинформационная структура. Впоследствии она поступает в гипоталамус и анализируется на рациональность и законченность. Сформированная матрица, удовлетворяющая этим критериям, принимается как новая программа и проявляется в виде фиксированной мысли. Это происходит не более чем в 2 % случаев. Законченность программы определяется ядрами гипоталамуса. Матрица, не удовлетворяющая требованиям рациональности, распадается. Второй вид блуждающих импульсов – импульсы поиска информации, или импульсы до востребования. Стимулом для их появления служит текущая информация. Такие импульсы циркулируют в мозге не постоянно, а генерируются в ситуациях, когда человеку нужно извлечь определённую информацию из арсенала памяти. Результатом их действия является закладка новой временной энергопрограммы в районе одной из стабилизирующих осей. Вначале программа довольно коротка. Дальнейшее же её формирование можно выразить так: когда человек думает о чём-то, в его памяти всплывает информация, относящаяся к данной теме. Это более характерно для состояния бодрствования. Но если человек достаточно «вжился» в проблему, то произведённый импульс до востребования продолжает существовать длительное время. Он может перейти из временного в относительно постоянный, и тогда циркулирование такого импульса и сбор им информации может продолжаться дни, месяцы и даже годы. В арсенал памяти информация, если она значима, поступает и в течение дня. Попадая в зону действия стабилизирующих осей, близкие к их текущей настройке информационные импульсы на материальных носителях начинают двигаться вдоль осей по направлению к ядерной зоне гипоталамуса. Достигнув области гипоталамуса, они, ударяясь о нижнюю выстилающую структуру, возвращаются к ядрам гипоталамуса или проходят ядерную зону. Такие цепочки, «замыкая» одну из шести стабилизирующих осей, становятся импульсами до востребования. Они имеют высокую степень свободы. Их дальнейший путь зависит как от собственной энергетической насыщенности, так и от состояния стабилизирующей оси. Если в данный момент энергетика импульса гомологична одной из шести стабилизирующих осей, то он переходит на эту ось. В противном случае имульс покидает гипоталамус и самостоятельно курсирует по подчерепным и арсенальным структурам. Взаимодействуя с имеющимися в арсенале программами, он собирает информацию, касающуюся поставленной задачи, т.е. согласующуюся с его кодом. Затем, попадая на стабилизирующую ось, он передаёт на неё собранную информацию и вместе с другими блуждающими импульсами участвует в формировании новой информационной матрицы будущей программы. Если информации для завершения матрицы недостаточно, то на стабилизирующей оси формируется новый импульс до востребования, который вновь засылается в арсенал памяти. Цикл существует до тех пор, пока формирующаяся матрица не достигнет законченности, что определяется ядрами гипоталамуса. При её достижении формируется биологическая структура, программа фиксируется сознанием и проявляется как фиксированная мысль. Если данный механизм работает, например, в процессе разговора, то сформированная матричная программа поступает в таламус и далее на выход: человек проговаривает собранную в арсенале памяти информацию. Приведём ещё пример. Поставлена установочная цель – сбор грибов. Реализуемая программа – временная. Поступая с таламуса на гипоталамус, она модулирует поле одной из шести стабилизирующих осей, образуя колебательный контур. Так формируется импульс до востребования, передающийся на арсенальные программы. Все программы, имеющие участки, резонирующие с пришедшим сигналом, переходят в возбуждённое состояние. Они выбрасывают на кору энергетические всплески, несущие информацию с этих участков. Энерговсплески воздействуют на сторожевые импульсы на материальных носителях, изменяя структуру их молекул. В нашем примере на сторожевых импульсах может быть записана информация о целесообразности сбора грибов, наличии тары и её объёме, условиях транспортировки и т.д. Данный цикл может повторяться неограниченное число раз. При этом к «рабочей» оси устремляется лавина сторожевых импульсов на материальных носителях, несущих результаты обработки информации арсенальными программами. Выстраиваясь вдоль оси и отвечая изменениями полевой структуры и, следовательно, поляризацией вещественной составляющей на излучаемое осью поле, они образуют матрицу временной программы. Матрица движется вдоль оси к гипоталамусу. Здесь она анализируется на законченность и в случае завершения осознается как фиксированная мысль. В противном случае процесс повторяется. Если формирование программной матрицы длится в течение 10–15 минут, то наступает торможение. Человек уже не видит грибов, думая о сопутствующих процессах. В этом случае необходимо отвлечься (или отдохнуть) и вновь запустить установку. Линия построения мыслей с просчитыванием какого-то варианта идёт равномерно, информационная же выборка всегда связана с постоянно циркулирующими блуждающими импульсами до востребования. Рассмотрим существующие взаимосвязи между стабилизирующими осями. Они выглядят в виде кольцевых энергетических цепей, не замкнутых до конца и имеющих разрыв до 1 мм (более полно информация рассмотрена в разделе «Арсенал памяти»). Назовем их энергомостами. Энергомосты отражают энергетические следы информационных процессов, но не единичных, а стабильно протекающих. Они упорядочивают движение блуждающих импульсов, то есть стабилизируют процесс. Генетически предопределено лишь то, что связи должны существовать, – у каждого индивида они расположены по-разному. Эти структуры формируются по энергоследу какого-либо устойчиво повторяющегося, стабильного акта логического мышления. Они не позволяют проходящим информационным цепям перехлестнуться – в этом их положительная роль. Отрицательное значение энергомостов состоит в том, что они препятствуют нестандартным решениям. При замыкании энергомоста процесс мышления (работа арсенала памяти) на некотором участке коры блокируется. Если связи «растягиваются», психика человека начинает вести себя максимально гибко при работе с ноовременными факторами, но это состояние граничит с шизофренией. Если данный эффект достигается в ходе движения по временному фактору с сознательным использованием процесса мышления, то опасности нарушения психики не существует. Для более детального рассмотрения изобразим оси схематично по принятой системе часового циферблата (рис. 2.9).
Оси не существуют автономно. С 1 до 5.30 все оси связаны энергомостами как с соседними осями, так и через одну, причём эти связи имеют довольно постоянный характер. Стабилизирующие оси связаны с биоэкраном, но не непосредственно, и эта связь не ярко выражена. С 3-й и 4-й осей на биоэкран поступает информация об интенсивности работы головного мозга («энергетический предохранитель»), сюда же поступают обратные сигналы на мозг. Не вся информация с биоэкрана проходит через оси на мозг – лишь только часть её в виде энергетического потенциала. Существующие связи между осями с 8 до 11 часов очень прочны и разрушаются только при нарушении морфологической целостности данной зоны.
Нельзя сказать, что энергомосты отвечают только за логику, хотя в этом они играют немаловажную роль, так как являются слепками логических цепей процесса мышления. Можно констатировать, что мосты отражают закономерные связи между накопленной в арсенале информацией, поэтому они строго индивидуальны. В области мозжечка больше связей между 3-й и 4-й, а также между 2-й и 5-й стабилизирующими осями. Здесь наблюдается несколько фиксированных энергомостов, которые зачастую замыкаются, образуя кольца. Такого рода «закольцовка» может носить кратковременный или долгосрочный характер. Долгосрочное замыкание энергомостов возникает при повреждении структур мозжечка (травмы) и при рассеянном склерозе. При этом закольцовка идёт двумя энергомостами в районе 2–3 часов и касается 3-й и 4-й стабилизирующих осей с последующим перемыканием коры больших полушарий на кору мозжечка (рис. 2.10). Кратковременное замыкание обусловливается психоэмоциональной деятельностью или информационной нагрузкой. Выделяются следующие возмущающие факторы: Особенно ярко это проявляется, когда психоэмоциональный всплеск длится очень короткий промежуток времени. Замыкание энергомоста при этом может длиться несколько часов. Возмущающие факторы могут быть связаны и с информационной перегрузкой: При описанных выше повреждающих факторах энергомосты замыкаются в районе 7–8 часов между 2-й и 5-й стабилизирующими осями. Замыкание энергомостов между 1-й и 6-й осями связано с существованием двух энерголучей, идущих от ромбовидной линзы в сагиттальном (спереди назад) направлении по коре. Ширина области формирования энергомостов в данном случае достаточно велика и варьирует от 1–2 до 8 часов по принятой шкале. На участке стабилизирующих осей с 1часа до 6.30 отвечают за логику приблизительно 20–30% энергомостов. Примерно 50% энергомостов отвечают за тонкое художественное восприятие и воспроизведение накопленных практических навыков. Оставшиеся 20–30% отражают в равной степени как практическое обучение подкорки, так и связь с биоэкраном и космосом. Таким образом, энергомосты – это не дендритные и аксонные связи, а энергетические структуры. Разрыв энергомостов в 1 мм есть своеобразная предохранительная вставка, которая позволяет системе работать достаточно гибко. Энергомосты позволяют тонко настраивать и регулировать процесс обработки информации, а в случае перегрузок – тормозить лавинообразные процессы. ПОДЧЕРЕПНОЙ ЭНЕРГОКОКОН
Подчерепной энергококон представляет собой энергетическую структуру, которая окутывает весь головной мозг между его корою и костью черепной коробки, не захватывая продолговатый мозг (рис. 2.11). Подчерепной энергококон выполняет дублирующие функции, перекликающиеся с работой стабилизирующих осей. Энергонасыщенность кокона велика. Она падает во время сна, когда механизмы обработки информации используются максимально и не требуют защиты от внешних воздействий. Поле энергококона стремится придать хаотическому движению блуждающих импульсов упорядоченность и стабильность. Это достигается путем дозированной подпитки блуждающих импульсов энергией или за счёт их стабилизации в период бодрствования. Поле имеет четыре основных пика спиралевидной формы, два из которых расположены в передней части мозга, в лобных долях, а два – в височной области. Как правило, они не соединяются с биоэкраном и не продуцируют никаких энергетических выбросов. Если смотреть из глубины мозга, эти образования закручиваются по часовой стрелке, образуя до 17 витков, не выходящих за пределы черепной коробки. В височной области энергоструктура подчерепного кокона входит в толщу костной ткани, хотя в других местах выстилает черепную коробку изнутри. По этой причине травмы височных областей чреваты тяжёлыми последствиями. Улиткообразные энергетические пики энергококона выполняют несколько функций: Работоспособность мозга прямо зависит от энергетической насыщенности подчерепного энергококона. Если информационная насыщенность блуждающих импульсов более или менее постоянна, то их энергетическая составляющая не всегда однородна. В результате значимая информация может не дойти до места назначения из-за слабой энергетики какого-либо фрагмента. СОН
Рассмотрим работу механизмов, характерных для сна, на примере его начальных стадий. При подготовке ко сну поэтапно протекают следующие процессы. 1. Программный анализ корковыми центрами состояния отдельных органов и систем, а также организма в целом. На этой стадии человек неосознанно и автоматически анализирует своё состояние. При отрицательном результате он либо устраняет имеющие место нарушения, либо «привыкает» к ним. Выбор варианта зависит от того, «настроена» ли данная программа на выздоровление или на дальнейшее развитие патологии. В первом случае устранение дефекта реализуется посредством гормонального механизма. Процесс протекает на стыке сознание-подсознание. И если работа всех систем удовлетворяет человека, а точнее, кору его мозга, то начинается второй этап. 2. Обработка максимально значимой информации. В этой фазе воспринимается и анализируется значимая информация, поступившая в течение дня, а также законченность программ. Человек пытается осмыслить основные события, прошедшие за день. Если какой-либо программе до завершения не хватает незначительного информационного блока, то мозг пытается спрогнозировать событие с учётом имеющейся информации или сразу отправить программу в арсенал памяти. В этот же период он может производить программирование событий на следующий день, то есть закладывать и выбирать те программы, которые могут понадобиться. За исключением первого этапа, все остальные могут проходить сознательно, неосознанно либо отсутствовать. На этом заканчивается подготовительная стадия и начинается период засыпания, сопровождающийся отключением многих систем организма. В зависимости от общего психоэмоционального настроя человека в данный момент процесс естественного засыпания может протекать по трём различным вариантам: Следующий период соответствует сну как таковому. Фаза глубокого сна наступает не постепенно, а сразу. Происходит отключение связи как с внешними анализаторами, так и с соматикой. Корковые механизмы переключаются полностью на восприятие и обработку информации дневного накопления. Моменты переключения коры на приём информации с матрицы дневного накопления могут сопровождаться двигательными реакциями (рефлекторное вздрагивание). В первой фазе сна подключаются комплексы, использующие биоэкран и максимально значимую информацию. При этом происходит непосредственное включение тех программ арсенала памяти, что заполняются в первой фазе медленного сна. Это основополагающее различие между сном и бодрствованием. В момент же бодрствования кора больших полушарий подключена непосредственно к зрительным и слуховым анализаторам. Сон крайне вариабелен, и время, необходимое для усвоения информации, может варьироваться у разных людей в широких границах. Время сна можно сократить без последствий для продолжительности жизни. Основополагающей здесь является кредовая позиция (жизненная установка). Она содержит в себе кольцевую программу с информационным кодом на определённый временной интервал сна, который каждый человек определяет себе сам. Процесс комплектования информации может идти не только ночью, но и в течение дня, например, в периоды отрешения и полудремоты. В это время происходит заполнение максимально значимой информацией созданных ранее программ арсенала памяти. В полном же объёме процесс протекает только во сне. В состоянии сна доминируют следующие факторы: Почти всегда (в 98%) сон начинается с медленной фазы, первая стадия которой длится от 2–3 до 10–15 минут. В этот промежуток считываются максимально значимые фрагменты информации – выступающие радикалы полинуклеотидной матрицы (без разрушения последней). На первой стадии медленного сна обрабатывается до 30% информации от всего дневного накопления. Наблюдается поляризация всей ткани гипоталамуса при участии ромбовидной линзы. На поляризованных участках группируются короткие транспортные структуры, которые могут образовывать небольшие цепи и в случае необходимости переносить объёмные разделы информации. Цепи принимают форму спиралей, закрученных против часовой стрелки по ходу движения, и транспортируются через ликвор в необходимые участки мозга. В основном, их конечная точка – арсенал памяти. Процесс его заполнения соответствует частотному диапазону до 35–70 кГц. Когда количество транспортных частиц уменьшается до 10–15%, начинается фаза быстрого сна. Это одна из основных причин смены фаз сна. Существует и ещё одна причина. Как только нарастает информационный поток, направленный на достройку программ арсенала памяти, начинает сказываться недостаток транспортировщиков, курсирующих по коре больших полушарий. Этот вид транспортных частиц выполняет распределительную функцию только во время сна. В состоянии бодрствования они располагаются на коре вдоль шести стабилизирующих осей. В первой стадии медленного сна до 60% от общего количества этих частиц используется для транспортных функций. Оставшиеся группируются по коре вдоль осей, обеспечивая стабильность работы головного мозга и исполняя роль энергетических предохранителей. Существуют и второстепенные причины, влияющие на смену фаз сна. Работа с нуждающимися в информации программами арсенала памяти ведёт к их активизации, усложняя процесс обработки. Активизированные программы накладывают свой отпечаток на компоновку дальнейшего быстрого сна. Кроме этого, необходимо время для построения программ на биоэкране, а также для управления органами и системами посредством гормональных изменений. Далее наступает новая фаза медленного сна, на которой вместе со считыванием информации происходит поэтапное разрушение полинуклеотидной матрицы. Частотные параметры следующих стадий медленного сна несколько выше первой и составляют 50–80 кГц. Так же выше и частотные параметры последующих стадий быстрого сна: на 5–7 кГц по отношению к первой. Таким образом, одна из причин чередования фаз сна – расход в фазах медленного сна биологических транспортных соединений. Пока длится фаза быстрого сна, идёт восстановление необходимого количества несущих субчастиц. На вопрос, почему мы видим сны, можно ответить так. Нужно учитывать, что, во-первых, происходит обработка информации с целью заполнения незавершённых программ. Она зависит от объёма информации и от наличия таких программ в арсенале памяти. Во-вторых, происходит энергетическая перестройка в биоэкране с подключением оболочечных структур. Это, в свою очередь, связано с фантомным, или оболочечным, перемещением во времени с последующим возвращением. Такое путешествие во времени приводит к появлению новой информации, которая также должна быть обработана. Так как подобная информация не поддерживается материальным носителем, то она недолговечна. Механизмы, обеспечивающие вспоминание временных путешествий и обработку подобных информационных комплексов, быстротечны. Максимальное время вспоминания – до 15 минут после пробуждения, хотя наиболее гомологичная программам арсенала памяти информация о временных путешествиях может оставаться довольно долго. Всё это можно отнести к сновиденческим комплексам. Некоторые фрагменты из сновиденческих комплексов переходят на стабилизирующие оси с последующей долгосрочной обработкой. Сохранение следов прохождения информации по определённым структурам больших полушарий позволяет хранить такого рода информацию на протяжении всей жизни. В период быстрого сна ассоциативная кора используется всего на 30%. Вокруг полинуклеотидной матрицы существует поле до 15–20 мм в диаметре, которое определяется её структурой. Если информация индифферентна и не затрагивает жизненное кредо данного человека, то положительная и отрицательная сторона этих полей совпадают, приводя к однородности. Если же доминирует одна из сторон – расшифровка матрицы проходит на негативном или позитивном фоне. В состоянии бодрствования это поле не оказывает существенного влияния на вегетативную систему, так как объём дневной обработки полинуклеотидной матрицы в 2–3 раза ниже, чем ночью. Подобные реакции чаще наблюдаются во время сна. Указанное поле воздействует на рецепторы гипофиза, который во время сна возбуждается за счёт бомбардировки его внешних слоёв субчастицами с ромбовидной линзы. Рецепторы гипофиза улавливают фоновое воздействие поля. Таким образом, информация, накапливаемая в течение дня, идёт на удовлетворение незаконченных программ арсенала памяти. В зависимости от её значимости она может «выбивать» из арсенала информационные фрагменты, которые проходят вдоль стабилизирующих осей длинными сериями с одновременной обработкой на зрительно-ассоциативных центрах коры больших полушарий и ромбовидной линзе. Это вызывает не только воспоминания в образах, но и их эмоциональную окрашенность. При прохождении через ромбовидную линзу «выбитые» информационные фрагменты захватываются полем полинуклеотидной матрицы и дополняют её. А так как в фазе быстрого сна происходит постоянная циркуляция информации, то к концу одного такого фрагмента программы арсенала памяти может пристраиваться другой. При этом наблюдается «скачка» сновидений, которая к окончанию сна нарастает. Космические Силы могут вклинивать не только свою информацию, но и производить её считывание. Процесс может затрагивать до 30% общего объёма информационных наработок и по времени продолжаться 10–15 минут в фазе конца быстрого и начала медленного сна. Кроме этого, некоторые информационные фрагменты могут «проигрываться» и расцениваться человеком как сон. Они проскакивают через 4–5 часов после начала сна или могут соответствовать фазе пробуждения. Если некоторые информационные фрагменты, снятые при обработке полинуклеотидной матрицы, имеют максимальную значимость для данного человека и затрагивают одну или несколько жизненно важных арсенальных программ, то наступает эффект резонанса. Человек как бы переживает эту ситуацию с подключением соматики. Образовавшийся информационный комплекс возбуждает кору больших полушарий. Это приводит к тому, что человек просыпается с ощущением всей гаммы последствий вегетативных реакций и сохраняет в памяти происшедшие во сне события. Такого рода «проблематичный» сон может проходить на фазе конца медленного и начала быстрого сна. Если человек в течение дня испытал сильную психоэмоциональную нагрузку, то во сне будут функционировать большие участки коры полушарий, и даже незначительная информация может вызвать описанный резонанс. При «проблематичном» сне частота колебаний в гипоталамусе может подниматься до отметки 120 кГц.
ГЛАВА 3. РОМБОВИДНАЯ ЛИНЗА СТРОЕНИЕ И ФУНКЦИИ
Рассмотрим механизмы декодирования информации, получаемой из внешней среды. Полинуклеотидная матрица дневного информационного накопления после объединения цепей и «опрессовки» направляется в конечную часть гипоталамуса и ориентируется по направлению к гипофизу. При этом своим острым концом она упирается в вещественно-энергетическую ромбовидную структуру. По своей геометрии она напоминает два конуса с общим основанием. Между конусами имеется промежуточный слой. Назовём данное образование ромбовидной линзой.
Промежуточный слой между двумя конусами состоит из диска, по обеим сторонам которого имеются прослойки. Молекулярная структура ромбовидной линзы представлена связанными между собой концентрическими ансамблями высокомолекулярного соединения. Основная функция ромбовидной линзы – обработка информации полинуклеотидной матрицы дневного накопления с последующим разрушением её материального носителя. При этом с матрицы снимается до 80% информации. Вещественная составляющая матрицы вызывает на линзе (при соприкосновении с её конусом) своеобразную реакцию. Вершина первого конуса не срезана, как в случае установочных линз таламуса, а слегка закруглена. Витки полинуклеотидных цепей как бы надеваются на этот закруглённый участок. При этом наблюдается поляризация микроструктур первого конуса, причём не только в точке касания. Волна поляризации проходит по всему первому конусу линзы до его основания. Как только первый виток с наименьшим диаметром касается вершины первого конуса, поляризуется его слой в зоне соприкосновения. Далее, по мере увеличения витков, фронт поляризации расширяется. Причём поляризационная волна в точности повторяет форму породившего её фрагмента спирали. Поляризация ромбовидной линзы происходит практически мгновенно с моментальным восстановлением первоначальной структуры. Вещественная часть матрицы, поляризуя верхние слои первого конуса, не проходит через структуру линзы. В состоянии сна ромбовидная линза поляризуется практически вся, так как полинуклеотидная матрица, раскручиваясь на линзе, увеличивает свои витки в диаметре до 1,5–2 см, возможно и до 3 см. Важнейший механизм прослойках и во втором конусе линзы. Здесь происходит считывание информации без участия субчастиц, имеющих нуклеотидную природу. Этим, однако, далеко не исчерпываются информационные процессы, протекающие в данной структуре. Большая часть из отколовшихся субчастиц в дальнейшем не разрушается. Разрушение их некоторой части связано с тем, что они несут доминирующую или значимую информацию, имеющую достаточно дублей, чтобы её не потерять. Разрушение других субчастиц связано с их энергетическим истощением. Перенося значимую информацию, субчастицы быстро теряют энергию, так как при считывании новому носителю максимально полно передаётся и энергетическая составляющая. Последующая репликация такой частицы возможна, однако, как правило, она не происходит, а её энергия отдается ромбовидной линзе. Отколовшиеся фрагменты полинуклеотидной матрицы не образуют цепи, но и не существуют хаотично. Субчастицы, химически не соединяясь друг с другом, за счёт полей формируют небольшие спирали. Они направляются во многие ядра головного мозга, попадая в его центральные участки, кору и подкорку. Формирование спиралей происходит по принципу информационного и энергетического сродства. Если информация чисто зрительная и относится к зрительно-ассоциативным звеньям, она направляется к центрам, «заинтересованным» в её получении. В дальнейшем информация из ядер поступает в подкорку и кору. На границе «ликвор – ядро» материальная составляющая откалывается, и информация в дальнейшем существует в виде энергоинформационного кванта, имеющего такую же структуру, что и исходный материальный носитель. Оставшаяся материальная часть фрагмента полностью разрушается и вместе с другими веществами поступает в ликвор для достройки и вовлечения в новые энергоинформационные процессы. Это ещё один путь движения информации для достройки программ арсенала памяти. Ядра имеют тракты, соединяющие их с арсеналом, которые могут передавать информацию либо перекрываться. Пройдя первый конус ромбовидной линзы, энергетическая поляризационная волна частично проходит во второй конус. Через первую прослойку на диск поступает только значимая и энергонасыщенная информация. Затем, проходя вторую прослойку, более жидкую по консистенции, чем первая, она поступает во второй конус. Если максимально значимую информацию принять за 1, то второго конуса линзы достигает информация с коэффициентом не ниже 0,4. Отсеянная на первой прослойке информация, не имеющая для человека большого значения, идёт на заполнение арсенальных программ или разрушается. Таким образом, первая прослойка и диск сортируют и компрессируют информацию. Пришедшая модулированная энергоинформационная волна поляризует второй конус, образуя спиральную волну, движущуюся по часовой стрелке. По мере прохождения по конусу витки спирали уменьшаются, а информация уплотняется. Этот процесс идёт на энергетическом уровне и более подробно будет рассмотрен далее. Информационно-энергетическая волна по мере прохождения оказывает влияние на гипофиз, что вызывает определённые вегетативные реакции и всплески со стороны всего организма в зависимости от значимости проходящей информации. Обработка накопленной матрицы происходит, как правило, во сне и лишь частично – в состоянии бодрствования, если поступившая информация жизненно необходима. Фрагментов такого рода за день поступает 2–3%. Они обгоняют нормальное течение «информационной реки» и, проходя второй конус, вызывают вегетативные реакции, от которых зависит жизнь человека. Хотя эта информация быстро разрушается, она успевает вызвать возмущения во всех отделах головного мозга, последующий синтез гормонов, обеспечивающих, в свою очередь, реакцию организма в целом. Во сне процесс разрушения полинуклеотидной матрицы ускоряется в три раза. При этом вся ромбовидная линза может поляризоваться. Иногда матрица не распадается на фрагменты, а расходится на две составляющие, образуя цепи, подобные тем, что формируются перед установочными линзами таламуса. Тогда они по каналам направляются на мозолистое тело. В 60% случаев так и происходит, если присутствует будничная незначимая информация. Если же она имеет кредовый характер, то такой механизм исключается. В норме на уровне продолговатого мозга и выше проходит энергетический слой, образующий своеобразное выпячивание. Этот слой служит покровом для ромбовидной линзы и окутывает полинуклеотидную матрицу дневного накопления. Данное поле осуществляет перенос информации кредового характера, причём поле существует не только в состоянии сна, но и при бодрствовании. Оно не имеет конкретной формы. Его можно представить в виде покрова, меняющего свое положение. Максимальная концентрация поля наблюдается в районах четвёртого мозгового желудочка и ромбовидной линзы. Его функционирование протекает параллельно процессам, идущим на стабилизирующих осях. В состоянии бодрствования происходит не только работа описанных ранее механизмов, но и локация окружающей среды биоэкраном с последующей передачей результатов в арсенал памяти. Кроме того, наблюдается совокупная деятельность по стабилизации информации при помощи стабилизирующих осей и подчерепного энергококона. |
||
|
|
Последнее изменение этой страницы: 2018-05-30; просмотров: 369. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |
 Поступающая в таламус по зрительным трактам информация весьма неоднородна для индивида по степени значимости. В ней содержатся как фрагменты стратегической, необходимой для оперативного арсенала, так и познавательной, различной степени важности, информации. На установочных линзах происходит разделение информационных фрагментов в зависимости от «степени заинтересованности» в них арсенальных структур. Та информация, которая проходит установочные линзы, менее значима. Она не является стратегической для индивида, а потому не влияет непосредственно на его жизненное кредо. Основная же часть полинуклеотидных цепей отсекается перед линзами и, предварительно их поляризовав, направляется в текущий арсенал памяти, как было описано выше.
Поступающая в таламус по зрительным трактам информация весьма неоднородна для индивида по степени значимости. В ней содержатся как фрагменты стратегической, необходимой для оперативного арсенала, так и познавательной, различной степени важности, информации. На установочных линзах происходит разделение информационных фрагментов в зависимости от «степени заинтересованности» в них арсенальных структур. Та информация, которая проходит установочные линзы, менее значима. Она не является стратегической для индивида, а потому не влияет непосредственно на его жизненное кредо. Основная же часть полинуклеотидных цепей отсекается перед линзами и, предварительно их поляризовав, направляется в текущий арсенал памяти, как было описано выше. Энергетическая составляющая прошедших информационных цепей дополнительно модулируется на линзах. Хотя самая важная информация считывается уже на подходе к линзам, ещё остаются блоки, значимость которых выше средней. Если информационный фрагмент по своим энергетическим параметрам близок текущему состоянию линз, то по пути следования он получает дополнительную энергетическую подпитку. Подобная информация с большей лёгкостью проникает через установочные линзы и быстрее обрабатывается.
Энергетическая составляющая прошедших информационных цепей дополнительно модулируется на линзах. Хотя самая важная информация считывается уже на подходе к линзам, ещё остаются блоки, значимость которых выше средней. Если информационный фрагмент по своим энергетическим параметрам близок текущему состоянию линз, то по пути следования он получает дополнительную энергетическую подпитку. Подобная информация с большей лёгкостью проникает через установочные линзы и быстрее обрабатывается. По мере продвижения остова ещё не достроенных полинуклеотидных цепей перемещается и фронт поляризации. Таламические субчастицы продолжают достраивать полинуклеотидную спираль, образуя своеобразное колено в противоположной стороне от входа в таламус. Достроившись на 80%, цепи разворачиваются на 180° и продолжают движение уже по нижним слоям долей таламуса в противоположную сторону. Проходя от колена расстояние до 1,5 см, они опускаются под углом в гипоталамус, одновременно сближаясь для соединения (рис. 2.6). Разворот цепей на 180° позволяет вести достройку верхней цепи за счёт наведённого поля нижнего участка таламуса и доукомплектовать эту часть матрицы. Таким образом происходит «опрессовка» полинуклеотидных цепей.
По мере продвижения остова ещё не достроенных полинуклеотидных цепей перемещается и фронт поляризации. Таламические субчастицы продолжают достраивать полинуклеотидную спираль, образуя своеобразное колено в противоположной стороне от входа в таламус. Достроившись на 80%, цепи разворачиваются на 180° и продолжают движение уже по нижним слоям долей таламуса в противоположную сторону. Проходя от колена расстояние до 1,5 см, они опускаются под углом в гипоталамус, одновременно сближаясь для соединения (рис. 2.6). Разворот цепей на 180° позволяет вести достройку верхней цепи за счёт наведённого поля нижнего участка таламуса и доукомплектовать эту часть матрицы. Таким образом происходит «опрессовка» полинуклеотидных цепей. полинуклеотидная матрица из двух спиралей – против часовой стрелки.
полинуклеотидная матрица из двух спиралей – против часовой стрелки.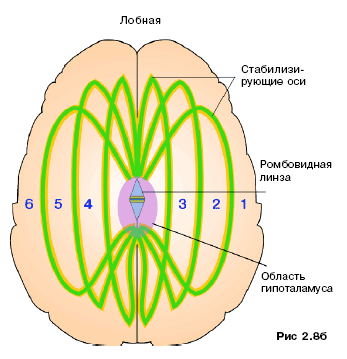 Отметим некоторые другие функции стабилизирующих осей. Во-первых, центральные оси (3-я и 4-я) энергетически дублируют фрагменты максимально значимой, не рутинной информации. Информационные блоки, которые «решаются» в данный момент, накладывают свой отпечаток на оси.
Отметим некоторые другие функции стабилизирующих осей. Во-первых, центральные оси (3-я и 4-я) энергетически дублируют фрагменты максимально значимой, не рутинной информации. Информационные блоки, которые «решаются» в данный момент, накладывают свой отпечаток на оси. Кроме 1-й и 6-й, стабилизирующие оси имеют высокую энергетическую насыщенность. Между лобной и затылочной частями осей также существует различие по энергонасыщенности. С 8 до 11 часов оси насыщены максимально. С 12 до 5.30 энергонапряжённость осей падает.
Кроме 1-й и 6-й, стабилизирующие оси имеют высокую энергетическую насыщенность. Между лобной и затылочной частями осей также существует различие по энергонасыщенности. С 8 до 11 часов оси насыщены максимально. С 12 до 5.30 энергонапряжённость осей падает. С 11 до 1 часа простирается «бессвязное» пространство между осями. Здесь могут существовать временные энергетические кольца – «неразвившиеся мосты». Они подобны миражам идей, витающим в воздухе. Временные кольца не являются энергетическими субстратами, рождающими идеи. Это отголоски процессов, происходящих на коре головного мозга при рождении идей.
С 11 до 1 часа простирается «бессвязное» пространство между осями. Здесь могут существовать временные энергетические кольца – «неразвившиеся мосты». Они подобны миражам идей, витающим в воздухе. Временные кольца не являются энергетическими субстратами, рождающими идеи. Это отголоски процессов, происходящих на коре головного мозга при рождении идей. Ромбовидная линза расположена под гипоталамусом, так что вершина первого конуса входит в гипоталамическую структуру, а вершина второго ориентирована по направлению к гипофизу (рис. 3.1).
Ромбовидная линза расположена под гипоталамусом, так что вершина первого конуса входит в гипоталамическую структуру, а вершина второго ориентирована по направлению к гипофизу (рис. 3.1).