|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
Ядерные и цитоплазматические мутацииСтр 1 из 2Следующая ⇒ Гибридное видообразование у растений Под гибридным видообразованием обычно подразумевают возникновение в потомстве от естественного гибрида новой линии, размножающейся в чистоте и изолированной от родительских видов и от своих сибсов в гибридной популяции. Эта новая линия должна преодолеть гибридную стерильность, и разрушение гибридов. Рекомбинационное видообразование Его можно определить как возникновение в потомстве видового гибрида с хромосомной стерильностью нового структурно-гомозиготного рекомбинанта, плодовитого при скрещиваниях с особями своей линии, но изолированного от других линий и от родительского вида преградой, создаваемой хромосомной стерильностью. Гибридное видообразование при участии внешних преград В некоторых группах растений межвидовые гибриды плодовиты и изоляция между видами обеспечивается главным образом внешними преградами. Экологическая и сезонная изоляция, а также изоляция, обусловленная строением цветка, — главные преграды, разделяющие виды. Морфологические, физиологические и поведенческие различия между видами, ведущие к возникновению таких преград, находятся, под контролем генов. У потомков естественных межвидовых гибридов, если они появляются, происходит расщепление по генным различиям и по соответствующим признакам, определяющим внешнюю изоляцию. Это создаёт возможность для возникновения продуктов межвидовой рекомбинации с новыми сочетаниями признаков, закладывающих основу новых, внешне изолированных субпопуляций. Если внешняя изоляция сохраняется и в дальнейшем, то из этих субпопуляций могут возникнуть новые виды гибридного происхождения. 
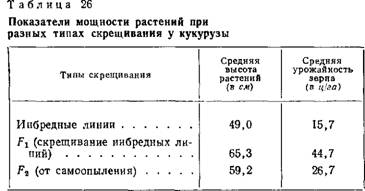
В селекции растений наиболее распространён метод гибридизации форм или сортов в пределах одного вида. С помощью этого метода создано большинство современных сортов сельскохозяйственных растений. Отдалённая гибридизация — более сложный и трудоёмкий метод получения гибридов. Основное препятствие получения отдалённых гибридов — несовместимость половых клеток скрещиваемых пар и стерильность гибридов первого и последующих поколений. Использование полиплоидии и возвратного скрещивания (беккросс) в отдельных случаях позволяет преодолеть не скрещиваемость пар и стерильность гибридов. Гибридизация ДНК Основная статья: Гибридизация ДНК Методы гибридизации ДНК состоят в смешивании одноцепочечных фрагментов ДНК, полученных от двух разных видов. Доля в смеси общей ДНК, которая воссоединяется, образуя двухцепочечные спирали, и скорость воссоединения служат мерами степени генетического родства между данными видами. Этот метод широко применяется зоологами, ботаниками и другими исследователями[1]. 6.Совр-оесост-е и достижение вселекции зарубежных стран. Она является теоретической основой селекции. На основе генетических исследований были разработаны методы получения гибридов кукурузы, подсолнечника, сахарной свеклы, огурца, а также гибридов и помесей животных, обладающих вследствие гетерозиса (гетерозис- это ускорение роста, увеличение размеров, повышение жизнеспособности и продуктивности гибридов первого поколения по сравнению с родительскими организмами) повышенной продуктивностью. Их дружба породила еще одно незаурядное произведение искусства: портрет Т. Г. Гнедич, написанный Акимовым - из лучших в созданной им портретной серии современников. Спектакль, поставленный и оформленный Акимовым в руководимом им ленинградском Театре Комедии, имел большой успех и продержался на сцене несколько лет. Первое представление, о котором шла речь в начале моего рассказа, завершилось триумфом Татьяны Гнедич. К тому времени тираж двух изданий "Дон Жуана" достиг ста пятидесяти тысяч, уже появилось новое издание книги К. И. Чуковского "Высокое искусство", в котором "Дон Жуан" оценивался как одно из лучших достижений современного поэтического перевода, уже вышла в свет и моя книга "Поэзия и перевод", где бегло излагалась история "Дон Жуана", причисленного мною к шедеврам переводческого искусства. И все же именно тот момент, когда поднялись с мест семьсот зрителей в зале Театра Комедии и все они единодушно благодарили вызванного на сцену "автора", стал наглядным апофеозом Татьяны Григорьевны Гнедич и ее удивительного творения Полиплоидия Полиплоидия (от греч. polýploos — многопутный, здесь — многократный и éidos — вид), кратное увеличение числа хромосом в клетках растений или животных. П. широко распространена в мире растений. Среди раздельнополых животных встречается редко, главным образом у аскарид и некоторых земноводных. Соматические клетки растений и животных, как правило, содержат двойное (диплоидное) число хромосом (2 n); одна из каждой пары гомологичных хромосом происходит от материнского, а другая — от отцовского организмов. В отличие от соматических, половые клетки имеют уменьшенное исходное (гаплоидное) число хромосом (n). В гаплоидных клетках каждая хромосома единична, не имеет парной себе гомологичной. Гаплоидное число хромосом в клетках организмов одного вида называется основным, или базовым, а совокупность генов, заключённую в таком гаплоидном наборе, — геномом. Гаплоидное число хромосом в половых клетках возникает вследствие редукции (уменьшения) вдвое числа хромосом в мейозе, а диплоидное число восстанавливается при оплодотворении. (Довольно часто у растений в диплоидной клетке бывают т. н. В-хромосомы, добавочные к какой-либо из хромосом.Роль их мало изучена, хотя у кукурузы, например, всегда имеются такие хромосомы.) Число хромосом у различных видов растений весьма разнообразно. Так, один из видов папоротника (Ophioglosumreticulata) имеет в диплоидном наборе 1260 хромосом, а у самого филогенетически развитого семейства сложноцветных вид Haplopappusgracilis имеет всего 2 хромосомы в гаплоидном наборе. При П. наблюдаются отклонения от диплоидного числа хромосом в соматических клетках и от гаплоидного — в половых. При П. могут возникать клетки, в которых каждая хромосома представлена трижды (3 n) —триплоидные, четырежды (4 n) — тетраплоидные, пять раз (5 n)— пентаплоидные и т.д. Организмы с соответственным кратным увеличением наборов хромосом — плоидности — в клетках называются триплоидами, тетраплоидами, пентаплоидами и т.д. или в целом — полиплоидами. Кратное увеличение числа хромосом в клетках может возникать под действием высокой или низкой температуры, ионизирующих излучений, химических веществ, а также в результате изменения физиологического состояния клетки. Механизм действия этих факторов сводится к нарушению расхождения хромосом в митозе или мейозе и образованию клеток с кратно увеличенным числом хромосом по сравнению с исходной клеткой. Из химических агентов, вызывающих нарушение правильного расхождения хромосом, наиболее эффективен алкалоид колхицин, препятствующий образованию нитейверетена деления клетки. (Воздействуя разбавленным раствором колхицина на семена и почки, легко получают экспериментальные полиплоиды у растений.) П. может возникать и вследствие эндомитоза — удвоения хромосом без деления ядра клетки. В случае нерасхождения хромосом в митозе (митотическая П.) образуются полиплоидные соматические клетки, при нерасхождении хромосом в мейозе (мейотическая П.) — половые клетки с измененным, чаще диплоидным, числом хромосом (т. н. нередуцированные гаметы). Слияние таких гамет даёт полиплоидную зиготу: тетраплоидную (4 n) — при слиянии двух диплоидных гамет, триплоидную (3 n) — при слиянии нередуцированной гаметы с нормальной гаплоидной и т.д. Возникновение клеток с числом хромосом 3-, 4-, 5-кратным (и более) гаплоидному набору, называется геномными мутациями, а получаемые формы — эуплоидными. Наряду с эуплоидией часто встречаетсяанеуплоидия, когда появляются клетки с изменением числа отдельных хромосом в геноме (например, у сахарного тростника, пшенично-ржаных гибридов и др.). Различают автополиплоидию — кратное увеличение числа хромосом одного и того же вида, и аллополиплоидию — кратное увеличение числа хромосом у гибридов при скрещивании разных видов (межвидовая и межродовая гибридизация). У полиплоидных форм растений нередко наблюдается гигантизм — увеличение размеров клеток и органов (листьев, цветков, плодов), а также повышение содержания ряда химических веществ, изменение сроков цветения и плодоношения. Эти особенности чаще наблюдаются у перекрёстноопыляющихся форм, чем у самоопылителей. Хозяйственно-полезные качества полиплоидов издавна привлекали внимание селекционеров, что привело к развёртыванию работ по искусственному получению полиплоидов, которые представляют важный источник изменчивости и могут быть использованы как исходный материал для селекции (например,. триплоидная сахарная свёкла, тетраплоидный клевер, редис и др.). Обычный недостаток автополиплоидов — низкая плодовитость. Однако после длительного отбора можно получить линии с достаточно высокой плодовитостью. Неплохие результаты даёт создание искусственных синтетических популяций, составленных из наиболее плодовитых линий автополиплоидов некоторых перекрёстноопыляющихся растений, например ржи. Не меньшее значение в селекции имеют и аллополиплоиды. Хромосомные наборы, входящие в состав аллополиплоидов, не одинаковы; они различаются набором содержащихся в них генов, а иногда формой и числом хромосом. При скрещивании растений разных родов, например ржи и пшеницы, возникает гибрид с гаплоидным набором ржи и гаплоидным набором пшеницы. Такой гибрид стерилен и лишь удвоение числа хромосом каждого растения, т. е. получение амфидиплоидов, может нормализовать мейоз и восстановить плодовитость. Аллополиплоидия может быть методом синтеза новых форм на основе гибридизации. Классический пример такого синтеза — получение Г. Д. Карпеченко рафанобрассики — гибрида редьки и капусты с 36 хромосомами (18 от редьки и 18 от капусты). Селекционерами (в СССР — В. Е. Писаревым, Н. В. Цициным, А. И. Державиным, А. Р. Жебраком и др.) аллополиплоиды получены у значительного числа видов растений. Большинство культурных растений, возделываемых человеком, — полиплоиды. П. имела огромное значение в эволюции дикорастущих и культурных растений (полагают, что около трети всех видов растений возникли за счёт П., хотя в некоторых группах, например у хвойных, грибов, это явление наблюдается редко), а также некоторых (преимущественно партеногенетических) групп животных. Доказательством роли П. в эволюции служат т. н. полиплоидные ряды, когда виды одного рода или семейства образуют эуплоидный ряд с увеличением числа хромосом, кратным основному гаплоидному (например, пшеница Triticummonococcum имеет 2n = 14 хромосом, Tr. turgidum и др. — 4n = 28, Tr. aestivum и др. —6n = 42). Полиплоидный ряд видов рода паслён (Solanum) представлен рядом форм с 12, 24, 36, 48, 60, 72 хромосомами. Среди партеногенетически размножающихся животных полиплоидные виды не менее часты, чем среди апомиктических растений (см. Апомиксис, Партеногенез). Советскому учёному Б. Л. Астаурову впервые удалось искусственно получить плодовитую полиплоидную форму (тетраплоид) из гибридов двух видов шелкопряда: Bombyxmori и В. mandarina. На основании этих работ им предложена гипотеза непрямого (через партеногенез и гибридизацию) происхождения раздельнополых полиплоидных видов животных в природе. 8.Мутагенез это внесение изменений в нуклеотидную последовательность ДНК (мутаций). Различают естественный (спонтанный) и искусственный (индуцированный) мутагенез. Естественный мутагенез Естественный, или спонтанный, мутагенез происходит вследствие воздействия на генетический материал живых организмов мутагенных факторов окружающей среды, таких какультрафиолет, радиация, химические мутагены. ]Мутационная теория Х. Де Фриза и С. И. Коржинского Мутационная теория составляет одну из основ генетики. Она зародилась вскоре после переоткрытия Т. Морганом законов Менделя в начале 20 столетия. Можно считать, что она почти одновременно зародилась в умах голландца Хуго Де Фриза[1] (1903) и отечественного ученого-ботаника С. И. Коржинского[2] (1899). Однако приоритет в первенстве и в большем совпадении изначальных положений принадлежит российскому ученому[3]. Признание основного эволюционного значения за дискретной изменчивостью и отрицание роли естественного отбора в теориях Коржинского и Де Фриза было связано с неразрешимостью в то время противоречия в эволюционном учении Ч. Дарвина между важной ролью мелких уклонений и их «поглощением» при скрещиваниях (см. кошмар Дженкина). Основные положения мутационной теории Коржинского-Де Фриза можно свести к следующим пунктам[4]: 1. Мутации внезапны, как дискретные изменения признаков 2. Новые формы устойчивы 3. В отличие от наследственных изменений, мутации не образуют непрерывных рядов, не группируются вокруг какого-либо среднего типа. Они являют собой качественные скачки изменений 4. Мутации проявляются по-разному и могут быть как полезными, так и вредными 5. Вероятность обнаружения мутаций зависит от числа исследуемых особей 6. Сходные мутации могут возникать неоднократно Механизм мутагенеза Последовательность событий приводящая к мутации (внутри хромосомы) выглядит следующим образом: · Происходит повреждение ДНК. · В случае, если повреждение произошло в незначащем (интрон) фрагменте ДНК, то мутации не происходит. · В случае если повреждение произошло в значащем фрагменте (экзон), и произошла корректная репарация ДНК, или вследствие вырожденности генетического кода не произошло нарушения, то мутации не происходит. · Только в случае такого повреждения ДНК, которое произошло в значащей части, которое не было корректно репарированно, которое изменило кодировку аминокислоты, или которое привело к выпадению части ДНК и соединению ДНК вновь в единую цепь — то оно приведет к мутации. Мутагенез на уровне генома также может быть связан с инверсиями, делециями, транслокациями, полиплоидией, и анеуплоидией, удвоением, утроением (множественной дупликацией) и т. д. некоторых хромосом. ]Точечные мутации Основная статья: Точечная мутация 1. Миссенс-мутация 2. Мутация сдвига рамки считывания 3. Нонсенс-мутация 4. Синонимическая сеймсенс-мутация. Хромосомные мутации 1. Инверсии 2. Реципрокные транслокации 3. Делеции 4. Дупликации и инсерционныетранслокации Геномные мутации 1. Анеуплоидия 2. Полиплоидия Ядерные и цитоплазматические мутации · Ядерные мутации — геномные, хромосомные, точечные. · Цитоплазмотические мутации — связанные с мутациями неядерных генов находящихся в митохондриальной ДНК и ДНК пластид — хлоропластов. ]Искусственный мутагенез Искусственный мутагенез широко используют для изучения белков и улучшения их свойств (направленной эволюции (англ.)). Ненаправленный мутагенез Методом ненаправленного мутагенеза в последовательность ДНК вносятся изменения с определенной вероятностью. Мутагенными факторами (мутагенами) могут быть различные химические и физические воздействия — мутагенные вещества, ультрафиолет, радиация. После получения мутантных организмов производят выявление (скрининг) и отбор тех, которые удовлетворяют цели мутагенеза. Ненаправленный мутагенез более трудоемок и его проведение оправдано, если разработана эффективная система скрининга мутантов. Направленный мутагенез В направленном (сайт-специфическом) мутагенезе изменения в ДНК вносятся в заранее известный сайт. Для этого синтезируют короткие одноцепочечные молекулы ДНК (праймеры),комплементарные целевой ДНК за исключением места мутации. Мутагенез по Кункелю Для бактериальной плазмиды (внехромосомной кольцевой ДНК) получают уридиновую матрицу, то есть такую же молекулу, в которой остатки тимина заменены на урацил. Праймер отжигают на матрице, проводят его достройку invitro с помощью полимеразы до кольцевой ДНК, комплементарной уридиновой матрице. Двухцепочечной гибридной ДНК трансформируют бактериальные клетки, внутри клетки уридиновая матрица разрушается как чужеродная, и на мутантной одноцепочеченой кольцевой ДНК достраивается вторая цепь. Эффективность такого способа мутагенеза менее 100 %. Мутагенез с помощью ПЦР Полимеразная цепная реакция позволяет проводить сайт-направленный мутагенез с использованием пары праймеров, несущих мутацию (рис. 1), а также случайный мутагенез. В последнем случае ошибки в последовательность ДНК вносятся полимеразой в условиях, понижающих ее специфичность 9.Аналитическая селекция.особенности селекционной работы, основанной на использовании местных сортов.Действие отбора не может выйти за определенные границы — оно должно прекратиться, когда будет получена чистая линия. До возникновения научной селекции улучшение возделываемых растений осуществлялось путем отбора лучших по внешним признакам особей. Еще в недалеком прошлом отбор был основным методом селекции. Исходным материалом для него обычно служили местные сорта, представляющие собой сложные популяции. При таком отборе селекционер выделяет только те генотипы, которые уже имеются в популяции. Вот почему селекцию того периода назвали аналитической. Отбор — один из главных факторов эволюции. В то время как мутации и рекомбинации создают генетическое разнообразие путем случайных изменений генетического кода (мутирование) или случайного расщепления в рамках законов Менделя, естественный отбор обеспечивает соответствие природных популяций с окружающей их средой. При этом негативный отбор приводит к устранению из популяции плохо приспособленных, а позитивный отбор обеспечивает сохранение хорошо приспособленных к сложившимся условиям внешней среды особей, что в итоге обусловливает поступательную и направленную эволюцию растений. Такое же важное значение имеет в современной селекции растений искусственный отбор. Это неотъемлемый элемент всех методов селекции, которые различаются между собой прежде всего способами создания исходного материала. Исходный материал может быть как естественного происхождения, так и получен путем скрещивания, полиплоидизации, мутагенеза. В то же время принципы отбора одинаковы для всех методов селекции. 9. Искусственный отбор, как уже упоминалось, применяется человеком во всех случаях, когда нужно выделить наиболее полезные растительные формы. После такого отбора каждое новое поколение должно быть лучше предыдущего. Искусственный отбор должен не только закреплять тот или иной полезный признак, но и развивать, усиливать его из поколения в поколение.
10.Методы отбора.творческая роль отбора. Селекция растений является наукой о создании новых сортов сельскохозяйственных культур. В ее задачу входит сбор, создание и изучение исходного материала, оценка полученных новых форм и образцов, испытание, районирование и внедрение в производство выведенных сортов. Главная цель селекционной работы заключается в создании новых сортов, способных давать более высокие урожаи, увеличивать валовые сборы продуктов растениеводства и улучшать их качество. Вся деятельность селекционных учреждений и селекционеров направлена на выполнение тех неотложных задач, которые стоят перед сельскохозяйственным производством по созданию достаточного количества продуктов питания для населения и сырья для легкой и пищевой промышленности. Под сортом понимается совокупность сходных по хозяйственно-биологическим свойствам и морфологическим признакам культурных растений, созданных и размноженных для возделывания в соответствующих природных и производственных условиях с целью повышения урожайности, качества продукции и экономической эффективности производства. Создаваемые человеком сорта предназначаются для получения высоких стабильных урожаев определенного вида продукции нужного качества. Растения, составляющие сорт, характеризуются общностью происхождения, имеют схожую генетическую основу и размножены из одного или
из нескольких исходных индивидуумов. Степень сходства растений, составляющих сорт, определяется как исходным селекционным материалом (гибриды, мутанты, гибридо-мутанты, полипоиды и др.), так и методами используемого отбора (индивидуальный, массовый, клоновый и др.). Сорт создается для определенных почвенно-климатических зон, где для него имеется возможность наиболее полной реализации потенциальной возможности генотипа. Сорта сельскохозяйственных культур по своему происхождению подразделяются на местные (созданные в результате действия естественного и искусственного отборов в определенной местности) и селекционные (созданные на основе научных методов селекции). В зависимости от способов выведения получают сорта-популяции, создаваемые путем массового отбора перекрестноопыляющихся (рожь, гречиха, кукуруза, свекла, клевер и др.) или самоопыляющихся растений; сорта-линии, получаемые путем индивидуального отбора растений самоопыляющихся культур (пшеница, ячмень, овес, горох, лен, люпин узколистный и др.), представляющие потомство, размноженное от одного растения. Сорта гибридного происхождения создаются путем скрещивания родительских форм с последующим отбором ценных растений для дальнейшего их размножения. Сорта-клоны, получаемые методом индивидуального отбора у вегетативно размножаемых культур (картофель, топинамбур, лук, чеснок, земляника). Гибриды в зависимости от способа получения подразделяются на простые, двойные, трехлинейные, межлинейные, сорто-линейные, линейно-сортовые. Для получения гибридов используют стерильные аналоги, фертильные аналоги закрепители стерильности и фертильные аналоги восстановители фертильности. Сорта и гибриды могут быть перспективными, районированными, дефицитными и стандартными. Перспективными признаются сорта и гибриды, которые успешно проходят государственное испытание, подтверждают свои преимущества, но требуют перед районированием прохождения производственного испытания и размножения в системе первичного семеноводства. Районированные сорта и гибриды заносятся в Государственный реестр охраняемых сортов и разрешаются для использования в производстве. Дефицитными являются районированные сорта и гибриды, по которым не полностью развернуто семеноводство и ощущается недостаток семян. Стандартом являются лучшие районированные сорта и гибриды, по которым в государственном испытании осуществляется сравнение всех испытываемых новых сортов на госсортоучастках (ГСУ) и государственных сортоиспытательных станциях (ГСС). К сорту, как средству сельскохозяйственного производства, предъявляются большие требования. В нем должна быть удачно совмещена высокая стабильная продуктивность с устойчивостью к неблагоприятным условиям производства, к болезням и вредителям. Он должен быть пластичным, т.е. обладать широким диапазоном нормы реакции, быть отзывчивым на вносимые удобрения и другие агротехнические приемы, иметь совершенную архитектонику. Пластичные сорта за счет особенностей генотипа и широты нормы реакции в государственном и производственном сортоиспытаниях дают более стабильные и высокие урожаи по годам на различных сортоучастках и сортоиспытательных станциях. На основании полученных данных такие сорта районируются на больших территориях и занимают обширные ареалы распространения. Примерами широкораспространенныхвысокопластичных сортов озимой ржи является сорт Вятка, Восход 1, Харьковская 60, озимой пшеницы Безостая 1, Мироновская 808, яровой пшеницы Лютесценс 62, Саратовская 29, Иволга, ярового ячменя Винер, Московский 121, Зазерский 85, Гонар, Гастинец, люпина Вайко, Быстрорастущий 4, Академический 1, картофеля Ранняя роза, Лорх, Приекульский ранний, Темп и другие. Продуктивность сорта любой сельскохозяйственной культуры зависит от его генотипа и условий окружающей среды. Особое значение имеют такие признаки, как зимостойкость, засухоустойчивость, устойчивость к болезням, вредителям и полеганию, дружность созревания, продолжительность вегетационного периода. Чем полнее соответствуют условия произрастания сорта его генотипу, тем продолжительнее сохраняются и стойко поддерживаются его типичность, чистота, высокие посевные качества и другие хозяйственно-биологические признаки и свойства. Вместе с тем известно, что условия произрастания культурных растений изменяются по годам, поэтому наблюдаются отклонения в реализации потенциальных возможностей сортов. Качество продукции определяется такими показателями как стекловидность, выполненность зерна, содержание белка и клейковины, выход муки или крупы у зерновых и крупяных культур, выход волокна у льна, крахмала у картофеля, сахара в свекле, кормопротеиновых единиц у трав и др. В любом научно-исследовательском учреждении селекция по созданию нового сорта начинается с разработки его модели (проекта), в основу которой берутся результаты тщательного изучения исходного материала и достижений селекции. Модель сорта включает перечень основных морфологических и хозяйственно-полезных признаков. В качестве эталона берется соответствующий лучший районированный сорт-стандарт с его параметрами. Во второй колонке модели указывается уровень показателей, который должен быть обеспечен в проектируемом сорте по урожайности и элементам ее структуры, устойчивости к болезням, вредителям, полеганию, осыпанию, неблагоприятным условиям погоды, качеству продукции, длине вегетационного периода и другим признакам. На протяжении всей истории селекции и особенно на ее современном научном этапе создано большое количество ценных сортов различных культур, изменивших во многих случаях облик и свойства своих предков до неузнаваемости. Яркими примерами этому могут служить достижения в селекции не только таких сравнительно новых культур, как сахарная свекла, подсолнечник, люпин, но и древних злаков - пшеницы, ржи, ячменя и других сельскохозяйственных растений. Подсолнечник, например, как масличное растение в полевой культуре, появился в России в 1816 году, но его посевы из-за сильной поражаемости заразихой и низкой урожайности не расширялись, а наоборот, после некоторого подъема к концу XIX века стали резко сокращаться. Начатая в 1912-1913 годах селекционная работа спасла подсолнечник как культуру и изменила коренным образом "цветок солнца". С 1977 года впервые в мировой практике районирован новый сорт подсолнечника Первенец, содержание олеиновой кислоты в масле которого достигает 70-75%, т.е. в два раза больше, чем у обычных сортов. Внедрение этого и подобных ему сортов в производство позволит полностью исключить импорт дорогостоящего оливкового масла, добываемого из плодов маслины в странах Средиземноморья. Под сортами селекции ВНИИМК им. В.С. Пустовойта в России и на Украине ежегодно занимаются площади в 4,4 млн. га и более 2 млн. га за рубежом и других странах почти всех континентов. Благодаря огромным успехам селекционеров краснодарские сорта подсолнечника даже на родине этой культуры, в странах американских континентов, не только получили широкое производственное распространение, но и являются основными источниками для селекционной работы в качестве исходного материала. Успех селекции подсолнечника заключается не только в повышении масличности его семян, но и в значительном повышении урожайности этой культуры. Значительно улучшены и такие хозяйственно-ценные свойства растения как устойчивость к болезням и вредителям, дружность созревания, пригодность к механизированной уборке и другие. Благодаря внедрению новых высокомасличных и высокоурожайных сортов сбор масла с 1 гектара достигает 2 тыс. и более кг/га, что почти в 5 раз больше по сравнению с 1940 годом. За счет этого ежегодно дополнительный сбор масла составляет сотни тысяч тонн подсолнечного масла. Второй весьма убедительный пример преобразования природы растений можно проследить на сахарной свекле. Впервые товарный сахар из этой культуры в Европе был получен в начале XIX столетия. Его содержание в то время в корнеплодах не превышало 6%, но благодаря начатой селекционной работе Луи Вильмореном в середине XIX века во Франции к 1888 году удалось повысить процент сахара в корнях до 10%, а через 10 лет до 15,2, в 1909 году лучшие сорта имели сахаристость до 18,4%. В настоящее время районированные сорта содержат в корнях до 20%сахара. Урожайность при этом увеличилась в несколько раз и достигла 45,0-50,0 т/га без орошения, что обеспечивает сбор сахара 7,5-9,0 т/га. Сорта сахарной свеклы Ганусовская односемянная 55, Белорусская односемянная 69, Кристалл, районированные в Беларуси, за ряд последних лет Государственного испытания на сортоучастках республики дают урожай корней до 50,3-68,0 т/га с выходом сахара до 9,3-10,6 т/га. Большим успехом в селекции сахарной свеклы является создание односемянных (одноростковых) сортов, что позволяет значительно уменьшить расход семян при посеве и максимально механизировать уход за плантациями этой культуры. Дальнейший прогресс в селекции этой культуры продолжается благодаря разработке новых селекционно-генетических методов с использованием полиплоидии и гибридизации сортов различной плоидности, при которой получают триплоидные гибридные семена в производственных масштабах, способные давать более высокие урожаи корнеплодов с повышенным содержанием сахара в них. Если сравнить эти достижения с теми несколькими центнерами урожая сахара, которые получали земледельцы от сахарной свеклы в начале XIX в., то можно сразу убедиться в огромной роли селекции и исключительно важном значении сорта. Результаты селекции люпина могут также служить ярким и выразительным примером превращения желтого, узколистного и белого люпинов, возделываемых на зеленое удобрение в высокобелковую ценную кормовую культуру, которая в условиях Белоруссии, Полесья Украины, нечерноземной зоне Российской Федерации и Прибалтийских республик имеет большое значение для решения проблемы белка. За 75 последних лет в результате селекционной работы ученых различных стран желтому люпину, например, приданы такие ценные признаки как безалкалоидность, белосемянность, нерастрескиваемость бобов, быстрые темпы первоначального роста, скороспелость, набухаемость семян без предварительной их скарификации, фузариозоустойчивость. За этот сравнительно короткий период времени у люпина, таким образом, были устранены многие признаки дикаря и создан обширный новый генофонд этой культуры для дальнейшей селекционной работы. Современные лучшие сорта люпина способны давать 1,5-2,0 тонны растительного полноценного по аминокислотному составу белка. Аналогичные примеры можно привести по картофелю, кукурузе, хлопчатнику и многим другим культурам. Нельзя обойти вниманием и выдающиеся успехи в селекции основной хлебной культуры пшеницы, история селекции которой насчитывает несколько тысячелетий. На всем ее протяжении люди методом отбора помогали эволюционному процессу, создали многочисленные местные сорта народной селекции. Однако они из-за целого ряда недостатков не в состоянии были обеспечить получение высоких урожаев. Среднегодовые урожаи озимой пшеницы даже на благоприятных землях Кубани за период 1901-1919 г.г. колебались от 0,6 до 1,3 т/га. И только благодаря усилию селекционной работы селекционерам в г. Краснодаре под руководством дважды Героя Социалистического Труда академика П.П. Лукьяненко удалось в конце 30-х годов создать сорт Краснодарская 622/2, превысивший наиболее распространенный в то время сорт Украинка на 10-15%. Этот небольшой скачек был очень важен для наращивания дальнейших успехов. Для создания устойчивых к полеганию и ржавчине сортов пшеницы был привлечен обширный исходный материал из американских и аргентинских сортов, в родословной которых участвовали сорта Англии, Голландии, Италии, Японии, Китая, Испании, Уругвая, США, России, Украины, Венгрии. При скрещивании аргентинского сорта Клейн 33 самериканскимКанред-Фулькастер 266287 были получены сорта Скороспелок 1, 2, 3 и 3б. Один из этой серии сорт Скороспелка 2 при скрещивании с украинским сортом Лютесценс 17 дал начало сортам нового типа, отличающихся устойчивостью к полеганию, высокой урожайностью и качеством зерна. Первый сорт этого типа Безостая 4 был районирован в 1955 году, а на его смену через 4 года пришел новый еще более ценный сорт Безостая 1, полученный методом индивидуального отбора из Безостой 4. Сорт Безостая 1 прославил советскую селекцию на весь мир и был справедливо назван шедевром мировой селекции. С внедрением в производство этих сортов урожайность озимой пшеницы на Кубани в 1963-1966 г.г. возросла почти в 3 раза по сравнению с дореволюционным периодом. 11. 12. 13. 14. Понятие о гетерозисе. В селекции животных и растений особое место занимает явление гибридной мощности, или гетерозиса, которое заключается в следующем. При скрещивании разных видов, рас, пород животных и сортов растений, а также инбредных линий гибриды Fi по ряду признаков и свойств часто превосходят исходные родительские организмы. Скрещивание гибридов Fi между собой ведет к затуханию этого эффекта в следующих поколениях. было показано, что скрещивание некоторых линий дает гибридные растения, более урожайные по зерну и вегетативной массе, чем исходные линии и сорта. В таблице 26 приведены опытные данные, показывающие низкую урожайность инбредных линий, значительное повышение урожайности в Fx и снижение ее в F2 при самоопылении растений. 15. Селе́кция (лат. selectio - выбирать) — наука о создании новых и улучшении существующих пород животных, сортов растений, штаммов микроорганизмов. Селекцией называют также отрасль сельского хозяйства, занимающуюся выведением новых сортов и гибридов сельскохозяйственных культур и пород животных. История Первоначально в основе селекции лежал искусственный отбор, когда человек отбирает растения или животных с интересующими его признаками. До XVI—XVII веков отбор происходил бессознательно: то есть человек, например, отбирал для посева лучшие, самые крупные семена пшеницы, не задумываясь о том, что он изменяет растения в нужном ему направлении. Только в последнее столетие человек, еще не зная законов генетики, стал использовать отбор сознательно или целенаправленно, скрещивая те растения, которые удовлетворяли его в наибольшей степени. Однако методом отбора человек не может получить принципиально новых свойств у разводимых организмов, так как при отборе можно выделить только те генотипы, которые уже существуют в популяции. Поэтому для получения новых пород и сортов животных и растений применяют гибридизацию, скрещивая растения с желательными признаками и в дальнейшем отбирая из потомства те особи, у которых полезные свойства выражены наиболее сильно. Например, один сорт пшеницы отличается прочным стволом и устойчив к полеганию, а сорт с тонкой соломиной не заражается стеблевой ржавчиной. При скрещивании растений из двух сортов в потомстве возникают различные комбинации признаков. Но отбирают именно те растения, которые одновременно имеют прочную соломину и не болеют стеблевой ржавчиной. Так создается новый сорт. Селекция и генетика В связи с развитием генетики, селекция получила новый импульс к развитию. Генная инженерия позволяет подвергать организмы целенаправленной модификации. Окончательно производится уже отбор лучших, но среди искусственно созданных генотипов. Селекция как наука оформилась лишь в последние десятилетия. В прошлом она была больше искусством, чем наукой. Навыки, знания и конкретный опыт, нередко засекреченный, были достоянием отдельных хозяйств, переходя от поколения к поколению. Только гению Дарвина удалось обобщить весь этот огромный и разрозненный опыт прошлого, выдвинув идею естественного и искусственного отбора как основного фактора эволюции наряду с наследственностью и изменчивость. Общие сведения Теоретической основой селекции является генетика, так как именно знание законов генетики позволяет целенаправленно управлять появлением мутаций, предсказывать результаты скрещивания, правильно проводить отбор гибридов. В результате применения знаний по генетике удалось создать более 10000 сортов пшеницы на основе нескольких исходных диких сортов, получить новые штаммы микроорганизмов, выделяющих пищевые белки, лекарственные вещества, витамины и т. п. К задачам современной селекции относится создание новых и улучшение уже существующих сортов растений, пород животных и штаммов микроорганизмов. Многолетняя селекционная работа позволила вывести много десятков пород домашних кур, отличающихся высокой яйценоскостью, большим весом, яркой окраской и т. п. А их единый предок — банкивская кура из Юго-Восточной Азии. На территории России не растут дикие представители рода крыжовник. Однако на основе вида крыжовник отклоненный, встречающийся на Западной Украине и Кавказе, получено более 300 сортов, многие из которых прекрасно плодоносят в России. Выдающийся генетик и селекционер академик Н. И. Вавилов писал, что селекционеры должны изучать и учитывать в своей работе следующие основные факторы: исходное сортовое и видовое разнообразие растений и животных; наследственную изменчивость; роль среды в развитии и проявлении нужных селекционеру признаков; закономерности наследования пригибридизации; формы искусственного отбора, направленные на выделение и закрепление необходимых признаков.
Селекция растений Основные методы селекции вообще и селекции растений в частности — отбор и гибридизация. Для перекрестноопыляемых растений применяют массовый отбор особей с желаемыми свойствами. В противном случае невозможно получить материал для дальнейшего скрещивания. Таким образом получают, например, новые сорта ржи. Эти сорта не являются генетически однородными. Если же желательно получение чистой линии — то есть генетически однородного сорта, то применяют индивидуальный отбор, при котором путем самоопыления получают потомство от одной единственной особи с желательными признаками. Таким методом были получены многие сорта пшеницы, капусты, и т. п. Для закрепления полезных наследственных свойств необходимо повысить гомозиготность нового сорта. Иногда для этого применяют самоопыление перекрестноопыляемых растений. При этом могут фенотипически проявиться неблагоприятные воздействия рецессивных генов. Основная причина этого — переход многих генов в гомозиготное состояние. У любого организма в генотипе постепенно накапливаются неблагоприятные мутантные гены. Они чаще всего рецессивны, и фенотипически не проявляются. Но при самоопылении они переходят в гомозиготное состояние, и возникает неблагоприятное наследственное изменение. В природе у самоопыляемых растений рецессивные мутантные гены быстро переходят в гомозиготное состояние, и такие растения погибают, выбраковываясь естественным отбором. Несмотря на неблагоприятные последствия самоопыления, его часто применяют у перекрестноопыляемых растений для получения гомозиготных («чистых») линий с нужными признаками. Это приводит к снижению урожайности. Однако затем проводят перекрестное опыление между разными самоопыляющимися линиями и в результате в ряде случаев получают высокоурожайные гибриды, обладающие нужными селекционеру свойствами. Это метод межлинейной гибридизации, при котором часто наблюдается эффект гетерозиса: гибриды первого поколения обладают высокой урожайностью и устойчивостью к неблагоприятным воздействиям. Гетерозис характерен для гибридов первого поколения, которые получаются при скрещивании не только разных линий, но и разных сортов и даже видов. Эффект гетерозиготной (или гибридной) мощности бывает сильным только в первом гибридном поколении, а в следующих поколениях постепенно снижается. Основная причина гетерозиса заключается в устранении в гибридах вредного проявления накопившихся рецессивных генов. Другая причина — объединение в гибридах доминантных генов родительских особей и взаимное усиление их эффектов. В селекции растений широко применяется экспериментальная полиплоидия, так как полиплоиды отличаются быстрым ростом, крупными размерами и высокой урожайностью. В сельскохозяйственной практике широко используются триплоидная сахарная свекла, четырехплоидный клевер, рожь и твердая пшеница, а также шестиплоидная мягкая пшеница. Получают искусственные полиплоиды при помощи химических веществ, которые разрушают веретено деления, в результате чего удвоившиеся хромосомы не могут разойтись, оставаясь в одном ядре. Одно из таких веществ — колхицин. Применение колхицина для получения искусственных полиплоидов является одним из примеров искусственногомутагенеза, применяемого при селекции растений. Путем искусственного мутагенеза и последующего отбора мутантов были получены новые высокоурожайные сорта ячменя и пшеницы. Этими же методами удалось получить новые штаммы грибов, выделяющие в 20 раз больше антибиотиков, чем исходные формы. Сейчас в мире культивируют более 250 сортов сельскохозяйственных растений, созданных при помощи физического и химического мутагенеза. Это сорта кукурузы, ячменя, сои, риса, томатов, подсолнечника, хлопчатника, декоративных растений. При создании новых сортов при помощи искусственного мутагенеза исследователи используют закон гомологических рядов Н. И. Вавилова. Организм, получивший в результате мутации новые свойства, называют мутантом. Большинство мутантов имеет сниженную жизнеспособность и отсеивается в процессе естественного отбора. Для эволюции или селекции новых пород и сортов необходимы те редкие особи, которые имеют благоприятные или нейтральные мутации. К одному из достижений современной генетики и селекции относится преодоление бесплодия межвидовых гибридов. Впервые это удалось сделать Г. Д. Карпеченко при получении капустно-редечного гибрида. В результате отдаленной гибридизации было получено новое культурное растение — тритикале — гибрид пшеницы с рожью. Отдаленная гибридизация широко применяется в плодоводстве. Выращивание сеянцев. В первый год развития сеянцы имеют только один недоразвитый лист. В мае — начале июня их высаживают в открытый грунт с площадью питания 50×30 см и выращивают здесь до начала цветения. 16. 17. Особенности селекции растений. Основными методами селекции растений является гибридизация и отбор, которые обычно применяют одновременно. Возможность получения значительного количества особей для селекционной работы позволяет использовать различные формы искусственного отбора: индивидуальный и массовый.
Прививки - особый способ искусственного объединения частей различных растений. Вы помните, что часть растения, которую прививают в другую, называют привоем, а растение, в которую прививают-подвоем. Прививка не является настоящей гибридизацией, ибо приводит лишь к ненаследственных изменений фенотипа такой объединенной растения, не меняя генотип ни с исходных организмов. Прививки применяют в разных целях: 18. Подготовка семян к посеву, наверно самый важный этап во всей посевной компании. Все дальнейшее зависит от того как вы подготовите семена и тут есть несколько советов. Подготовка семян к посеву начинается с отбора наиболее крупных, полновесных семян. Такие семена, как правило, дают наиболее высокий урожай. Для отбора все семена высыпают в 3-5 % раствор (30-50 грамм на 1 литр воды) поваренной соли. Семена аккуратно высыпают небольшими порциями, помешивая. Через 3-5 минут легкие, всплывшие на поверхность семена удаляют. Полновесные семена оседают на дно посуды, их тщательно промывают под струей воды и просушивают, рассыпая тонким слоем и систематически помешивая. После сортировки семена проверяют на всхожесть. Для этого нужно всего лишь на дно плоской тарелки или блюдца расстелить сложенную в 3 — 4 слоя марлю, газету или ткань, увлажнить и разложить семена. Сверху семена покрывают этой же тканью, смачивают теплой водой, закрывают тарелкой или блюдцем и ставят в теплое место (20-22°). Следят, чтобы ткань была постоянно влажной. Хорошо под слой ткани, на которую укладываются для проращивания семена, положить слой ошпаренных кипятком опилок. Сверху на слой ткани, покрывающей семена, также можно насыпать опилки слоем в 1,5-2,0 см. 19. 20. Методы оценки селекционного материала. В процессе селекции материал оценивают по хозяйственным и биологическим свойствам, являющимся объектом селекции. Различают: 1) полевые методы оценки, когда оценка проводится непосредственно в полевых условиях; 2) лабораторные методы оценки, куда относятся оценки по качеству урожая с применением необходимых для этого анализов, а также оценки на устойчивость к неблагоприятным факторам в специальных камерах или установках; 3) лабораторно-полевые методы, когда подготовка материала к оценке ведется в одних условиях (в лаборатории или в поле), а затем подготовленный материал помещается для окончательной оценки в другие условия (поле или лаборатория). Например, предпосевная закалка семян к засухе ведется в лаборатории, а закаленные семена высевают затем в поле. Закалку озимых против мороза проводят в полевых условиях, а затем молодые растения переносят для испытания в холодные камеры. Методы оценки должны быть по возможности простыми, но точными, основанными на использовании малых количеств материала, и высокопроизводительными. Методы оценки можно также разделить на прямые, когда непосредственно определяют степень выраженности интересующего нас признака, и косвенные, когда об этом признаке судят по другим, связанным с ним признакам... Так, устойчивость растений к морозу можно оценить, подвергая их замораживанию при определенных температурах (прямой метод) или же определяя в них содержание Сахаров после закалки (косвенный метод). Существуют еще провокационные методы, когда искусственно создают среду для выявления отношения растений к неблагоприятным для них условиям (морозу, засухе, болезням), чтобы затем выбраковать пострадавшие растения. 21. 22. Сортовой контроль, установление принадлежности растений и семян к тому или иному сорту растений(или гибриду) и определение их сортовой чистоты. Для С. к. применяют полевую (растения осматривают в поле на корню или отбирают и анализируют апробационный сноп, образец) и амбарную (семена апробируют в семенохранилище) апробацию, грунтовой контроль (средний образец семян высевают на делянках и по растениям проверяют сортовые признаки), лабораторный контроль (изучают семена, их проростки и всходы в лаборатории, используют органолептические, химические и физические методы, что позволяет выявить признаки сорта в любое до посева время). Входит в систему семенного контроля 23. |
||
|
|
Последнее изменение этой страницы: 2018-05-30; просмотров: 1341. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |