|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
Хромосомная теория Т. Моргана1. Гены находятся в хромосомах. Каждый ген занимает в хромосоме строго определённое место (локус). Гены в хромосомах расположены линейно. 2. Гены, которые находятся в одной хромосоме, представляют собой группу сцепления генов. Число групп сцепления равно гаплоидному набору хромосом. 3. Сцепление генов может нарушаться в результате кроссинговера (перекреста хромосом), в процессе которого между гомологичными хромосомами происходит обмен одним или несколькими аллельными генами. 4. Расстояние между генами в хромосоме прямо пропорционально частоте кроссинговера между ними. Сцепленное наследование – наследование признаков, за которые отвечают гены, расположенные в одной хромосоме. При полном сцеплении кроссинговер между генами по каким-либо причинам невозможен, или гены близко расположены друг к другу и вероятность кроссинговера между ними настолько мала, что им можно пренебречь. При частичном сцеплении между генами возможен кроссинговер. Кроссинговер происходит не всегда, поэтому кроссоверных (рекомбинантных) особей значительно меньше, чем количество основных особей. Это может служить критерием для определения сцепления генов, находятся ли гены в одной хромосоме или в разных парах гомологичных хромосом. На основании анализа частоты кроссинговера определяют расстояние между генами и составляют генетические карты хромосом. Расстояние между генами выражают в процентах кроссинговера или морганидах (1 морганида соответствует 1% кроссинговера). Генетика пола Различают несколько вариантов определения пола: 1. Пол определяется составом половых хромосом:  - у млекопитающих женский пол – гомогаметный (несет одинаковые половые хромосомы ХХ и образует один тип гамет), мужской пол - гетерогаметный (несет разные половые хромосомы ХУ и образует 2 типа гамет); - у птиц, бабочек гомогаметный пол мужской (ZZ), гетерогаметный – женский пол (ZW); 2. Пол определяется числом половых хромосом. Например, у кузнечиков самки - ХХ, а самцы – X0. 3. Пол определяется соотношением числа Х-хромосом и числа наборов аутосом (такое отношение называется половым индексом). Такой тип определения пола характерен для дрозофилы. Если половой индекс равен 1 (2Х:2А), то развивается самка, если он равен 0,5 (1Х:2А), - развивается самец. Y-хромосома у дрозофил не определяет пола, однако в ней находятся гены фертильности самцов. Самцы Х0 стерильны. 4. Гапло-диплоидный тип определения пола. У пчел, муравьев, ос нет половых хромосом. Самки развиваются из оплодотворенных яиц и имеют диплоидный набор хромосом, а самцы - из неоплодотворенных яиц и имеют гаплоидный набор хромосом. 5. Пол определяется внешними условиями (например, у некоторых рептилий пол определяется температурой развития яиц).
Изменчивость 1. Ненаследственная (фенотипическая или модификационная) изменчивость – изменчивость, не связанная с изменением генотипа и возникающая под влиянием условий среды. Пределы модификационной изменчивости называют нормой реакции. Норма реакции признака определяется генотипом. Различают широкую норму реакции (признак изменяется в широком диапазоне) и узкую. Свойства модификационной изменчивости: 1) не наследуется; 2) имеет приспособительный характер; 3) имеет массовый (групповой) характер, т.е. проявляется у группы особей, находящихся в одинаковых условиях; 4) адекватна изменениям условий среды и предсказуема (Ч.Дарвин называл модификационную изменчивость определенной); 5) носит постепенный характер; 6) обратима, т.е. модификации исчезают, если прекращается действие вызвавшего их фактора. 2. Наследственная изменчивость связана с изменениями генотипа. а) комбинативная изменчивость является результатом перекомбинации генов у потомков, возникает при половом размножении. Механизмы комбинативной изменчивости: · кроссинговер (в профазе I мейоза при гаметогенезе); · независимое расхождение гомологичных хромосом (в анафазу I мейоза при гаметогенезе); · случайное слияние гамет при оплодотворении. Значение: способствует увеличению генетического разнообразия потомства; лежит в основе гибридологического метода в генетике и селекции, позволяет получать организмы с необходимыми человеку комбинациями признаков. б) мутационная изменчивость – вызывается мутациями – непредсказуемыми скачкообразными изменениями генотипа. Термин предложен Г.де Фризом в 1901г. Факторы, вызывающие мутацию, называются мутагены.Различают: - физические мутагены (ионизирующее излучение, ультрафиолетовые лучи); - химические (формалин, аналоги азотистых оснований, соли тяжелых металлов, гербициды); - биологические (ДНК вирусов, гельминты и продукты их жизнедеятельности). Классификации мутаций: 1. По уровню организации генетического материала, на котором возникла мутация: генные, хромосомные и геномные. 2. По влиянию на жизнеспособность: летальные (приводят к гибели организма), полулетальные (снижают жизнеспособность, организм обычно не доживает до репродуктивного возраста), нейтральные (не влияют на жизнеспособность), благоприятные. 3. По проявлению в гетерозиготе: доминантные и рецессивные. 4. По типу клеток, в которых возникает мутация: соматические (возникают в клетках тела и при половом размножении не наследуются) и генеративные (возникают в половых клетках). 5. По локализации в клетке: ядерные (в ДНК ядра) и цитоплазматические (в ДНК митохондрий и пластид). 6. По фенотипическому проявлению: биохимические, физиологические морфологические. 7. По причине, вызывающей мутацию: спонтанные (возникают самопроизвольно), и индуцированные (возникают при экспериментальном воздействии на генетический материал).
Генные мутации - изменения числа или последовательностинуклеотидов в структуре ДНК (вставки, выпадения, перемещения, замена нуклеотидов) в пределах отдельных генов. Могут возникать при репликации ДНК и приводят к изменению количества или качества соответствующих белков. Генные мутации в зависимости от механизма и последствий мутаций можно разделить на две группы: 1. Мутации со сдвигом рамки считывания информации (вставка, выпадение, перемещение одного или нескольких нуклеотидов). В связи с тем, что генетический код не имеет знаков препинания, меняется состав всех триплетов после места мутации, что приводит к замене многих аминокислот и синтезу белка с измененными свойствами. 2. Мутации без сдвига рамки считывания информации (замена нуклеотида). При этом меняется состав одного триплета, что приводит к изменению только одной аминокислоты в белке. Такая замена не всегда существенно сказывается на изменении свойств белка (особенно, если новая аминокислота по свойствам близка к исходной). Однако в ряде случаев, замена даже одной аминокислоты может привести к серьезным последствиям. Так, замена в шестом положении глутаминовой кислоты β-цепи гемоглобина на валин приводит к развитию тяжелого заболевания – серповидно-клеточной анемии. Генные мутации играют большую роль в эволюции. Они создают резерв наследственной изменчивости и представляют материал для естественного отбора. Хромосомные мутации (аберрации) связаны с изменением структуры хромосом. Могут возникать при нарушении конъюгации хромосом и кроссинговера в мейозе. 1. Внутрихромосомные перестройки а) делеции - утрата участка хромосомы. Делеции являются результатом разрывов хромосом. б) дупликации - локальное удвоение определенного участка хромосомы. в) инверсии - поворот на 180° участка хромосомы. Если утраченные или удвоенные при кроссинговере участки хромосом очень малы, они не влияют на жизнеспособность гамет и образуемых при их слиянии зигот. Инверсии в хромосомах человека приводят к нарушению гаметогенеза. 2. Межхромосомные перестройки (транслокации). а) реципрокные транслокации – взаимный обмен, т.е. обмен фрагментами между двумя негомологичными хромосомами; б) нереципрокные транслокации – перемещение участка одной хромосомы на другую, негомологичную ей; в) робертсоновские транслокации – слияние негомологичных акроцентрических хромосом в области их центромер с образованием одной метацентрической хромосомы. При этом число хромосом в кариотипе уменьшается на одну, так как обычно одна из центромер и короткие плечи двух акроцентрических хромосом утрачиваются. г) инсерции – перемещение фрагмента одной хромосомы внутрь другой. Геномные мутации связаны с изменением числа хромосом. 1. Эуплоидия – изменение числа хромосом, кратное гаплоидному набору. а) гаплоидия – уменьшение числа хромосом, кратное гаплоидному набору. Клетки гаплоидов содержат одинарный набор хромосом (n). Гаплоидные организмы обычно мельче диплоидных; менее жизнеспособны, т.к. у них проявляются рецессивные гены с неблагоприятным действием; стерильны. Экспериментальное получение гаплоидных организмов (растений) позволяет изучать проявление рецессивных мутаций, а также получать чистые линии растений путем перевода гаплоидов в диплоидное состояние. б) полиплоидия – увеличение числа хромосом, кратное гаплоидному набору. Различают: · автополиплоидию - повторение одного и тот же хромосомного набора, часто встречается у растений; · аллополиплоидию - повторение разных хромосомных наборов. В природе встречается редко, возникает при межвидовой гибридизации, чтобы гибриды смогли дать потомство необходима полиплоидизация. Так, Г.Д. Карпеченко был получен плодовитый гибрид капусты и редьки. Полиплоидия может возникнуть в результате нарушения расхождения хромосом в митозе; слияния соматических клеток; нарушения мейоза, в результате чего возникают гаметы с нередуцированным числом хромосом. 2. Анэуплоидия (гетероплоидия) - изменение числа хромосом, некратные гаплоидному набору. а) нуллисомия - отсутствие обоих гомологов какой-либо одной паре хромосом (общее число хромосом в такой клетке равно 2n-2). б) моносомия - утрата одного из гомологов по одной или большему числу пар хромосом (2n -1). Наиболее известная моносомия у человека - синдром Шерешевского-Тернера (женщины с кариотипом 45, Х0). в) полисомия - избыточное число гомологичных хромосом в одной или нескольких парах гомологичных хромосомом. Наличие одной дополнительной гомологичной хромосомы приводит к трисомии (2n +1), двух - к тетрасомии (2n +2). Наиболее распространенные синдромы с трисомией по отдельным аутосомам: синдром Патау (47,+13), синдром Эдвардса (47,+18), синдром Дауна (47,+21). Описана трисомия по половым хромосомам - синдром Клайнфельтера (47, ХХУ), а также его варианты с тетрасомией и пентасомией. Возникновение анэуплоидий может быть связано с нерасхождением отдельных хромосом в первом делении мейоза у одного из родителей или с нарушением второго деления мейоза.
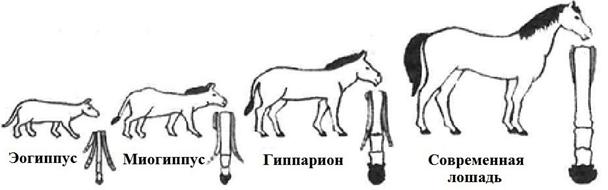
Эволюция Доказательства эволюции 1.Палеонтологические доказательства основаны на изучении ископаемых форм организмов. а) ископаемые переходные формы – формы организмов, занимающие промежуточное положение между крупными систематическими группами. Например, кистеперая рыба Эустеноптерон – переходная форма между рыбами и наземными четвероногими земноводными, археоптерикс – переходная форма между рептилиями и птицами, лицено́пс - переходная форма между рептилиями и млекопитающими. б) филогенетические ряды – ряд ископаемых форм, последовательно связанных друг с другом в процессе эволюции. Формы в филогенетических рядах близки по основным и частным деталям строения, связаны генеалогически. Например, известны филогенетические ряды лошадей (рис. 27), носорогов, слонов и др.
Рис. 27. Филогенетический ряд лошади
2. Сравнительно-анатомические(морфологические)доказательства основываются на изучении сходства и различий в строении организмов. Сходство внутреннего строения организмов может свидетельствовать об их родстве. а) гомологичные органы – органы, сходные по происхождению и строению, но выполняющие разные функции (например, передние конечности позвоночных - ласты кита, лапы крота, крылья птицы, руки человека). Гомологичные органы возникают вследствие расхождения признаков (дивергенции) в разных условиях обитания организмов. Аналогичные органы – органы, выполняющие одинаковые функции, но различающиеся происхождением и строением (например, крыло птицы и бабочки, глаз позвоночных животных и головоногих моллюсков, роющие конечности крота и насекомого медведки). Аналогичные органы возникают вследствие схождения (конвергенции) признаков и не свидетельствуют о родстве между организмами (рис. 28)
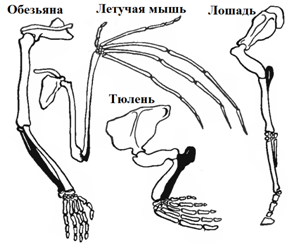
А Б Рис. 28. Гомологичные органы позвоночных животных (А), аналогичные конечности крота и медведки (Б)
б) рудименты – недоразвитые органы, утратившие в ходе эволюции свое значение (третье веко, копчик, червеобразный отросток, зубы мудрости, мышцы, двигающие ушную раковину). Атавизмы – проявление признаков, свойственных предкам, но утраченных в процессе эволюции (хвостовой придаток, сильный волосяной покров, многососковость у человека, развитие настоящих боковых пальцев у лошади). Проявляются у отдельных организмов, показывают историческую взаимосвязь вымерших и существующих форм. в) Переходные формы – формы, объединяющие в своем строении признаки разных типов организации. Занимают промежуточное систематическое положение. Например, эвглена зеленая сочетает признаки животных и растений. 3. Эмбриологические доказательства основаны на сопоставлении зародышей разных таксонов на определенных стадиях эмбрионального развития. а) закон зародышевого сходства К.М.Бэра: чем более ранние стадии индивидуального развития исследуются, тем больше сходства обнаруживается между различными организмами. б) биогенетический закон Геккеля-Мюллера: онтогенез – краткое повторение филогенеза. Позже А.Н. Северцов доказал, что изменения исторического развития обусловлены изменениями зародышевого развития (теория филэмбриогенезов). 4. Биогеографическиедоказательства основаны на сравнении фауны и флоры разных континентов, изучении распространения близких форм, изучении островных и реликтовых форм. Реликты – отдельные виды с комплексом признаков, характерных для древних вымерших организмов (гаттерия, кистеперая рыба латимерия, гинкго). 5. Молекулярно-генетическиедоказательства заключаются в сравнительном изучении кариотипов, последовательностей нуклеотидов ДНК, анализе белкового состава крови. |
||||||
|
|
Последнее изменение этой страницы: 2018-05-29; просмотров: 378. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |