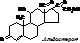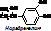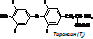|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
Гормоны паращитовидных железКальцитриол. Строение Представляет собой производное витамина D и относится к стероидам. Синтез Образующийся в коже под действием ультрафиолета и поступающие с пищей холекальциферол (витамин D3) и эргокальциферол (витамин D2) гидроксилируются в печени по С25 и в почках по С1. В результате формируется1,25-диоксикальциферол(кальцитриол). Регуляция синтеза и секреции Активируют: Гипокальциемия повышает гидроксилирование по С1 в почках.Уменьшают: Избыток кальцитриола подавляет гидроксилирование по С1 в почках.Механизм действия Цитозольный. Мишени и эффекты Эффект кальцитриола заключается в увеличении концентрации кальция и фосфора в крови: oв кишечнике индуцирует синтез белков, отвечающих за всасывание кальция и фосфатов, o впочках повышает реабсорбцию кальция и фосфатов,oвкостной ткани усиливает резорбцию кальция.
ПАРАТИРЕОИДНЫЙ ГОРМОН Строение Представляет собой пептид из 84 аминокислот с молекулярной массой 9,5 кДа. Синтез Идет в паращитовидных железах. Реакции синтеза гормона высоко активны. Регуляция синтеза и секрецииВысокие концентрации кальция активируют кальций-чувствительнуюпротеазу, гидролизующую один из предшественников гормона, и тем самым снижается образование паратиреоидного гормона. Механизм действия Аденилатциклазный. Мишени и эффекты Эффект паратиреоидного гормона заключается в увеличении концентрации кальция и снижении концентрации фосфора в крови. Это достигается тремя способами: Костная ткань o при высоком уровне гормона активируютсяостеокластыи происходит деструкция костной ткани. При низких концентрациях активируется перестройка кости и остеогенез. Почкиo увеличивается реабсорбциякальцияи магния,  oуменьшается реабсорбция фосфатов, аминокислот, карбонатов, натрия, хлоридов, сульфатов. o также гормон стимулирует образование кальцитриола (гидроксилирование по С1). Кишечник o при участии кальцитриола усиливается всасывание кальция и фосфатов. Гипофункция Возникает при случайном удалении при операциях на щитовидной железе, аутоиммунной деструкции ткани желез. Возникающая гипокальциемия и гиперфосфатемия проявляется в виде высокой нервно-мышечнойвозбудимости, судорог, тетании. При резком снижении кальция возникает дыхательный паралич, ларингоспазм. Гиперфункция Первичный гиперпаратиреоз возникает при аденоме желез. Нарастающая гиперкальциемия вызывает повреждение почек, мочекаменную болезнь Вторичный гиперпаратиреоз является результатом почечной недостаточности, при которой происходит нарушение образования кальцитриола, снижение концентрации кальция и компенсаторное возрастание синтеза паратиреоидного гормона. КАЛЬЦИТОНИн Строение Представляет собой пептид, включающий 32 аминокислоты с молекулярной массой 3,6 кДа. Синтез Осуществляется в парафолликулярных клетках щитовидной железы. Регуляция синтеза и секреции Активируют:ионы кальция, глюкагон. Механизм действия Аденилатциклазный Мишени и эффекты Эффект кальцитонина заключается в уменьшении концентрации кальция и фосфора в крови. Вкостной ткани подавляет активность остеокластов, что улучшает вход кальция и фосфатов в кость. Впочках подавляет реабсорбцию Ca, P, Na, K, Mg. Патология не отмечено.
Регуляция водно-солевого обмена. Роль вазопрессина, альдостерона и ренин-ангиотензиновой системОсновными параметрами водно-солевого гомеостаза являются осмотическое давление, рН и объем внутриклеточной и внеклеточной жидкости. Изменение этих параметров может привести к изменению артериального давления, ацидозу или алкалозу, дегидратации и отекам. Основными гормонами, участвующими в регуляции водно-солевого баланса, являютсяАДГ, альдостерон и предсердный натрий-уретический фактор (ПНФ). АДГ, или вазопрессин, — пептид, содержащий 9 аминокислот, соединенных одним дисульфидным мостиком. Синтезируется в виде прогормона в гипоталамусе, затем переносится в нервные окончания задней доли гипофиза, из которых секретируется в кровоток при соответствующей стимуляции. Перемещение по аксону связано со специфическим белком-переносчиком (нейрофизином) Стимулом, вызывающим секрецию АДГ, служит повышение концентрации ионов натрия и увеличение осмотического давления внеклеточной жидкости. Наиболее важные клетки-мишени для АДГ - клетки дистальных канальцев и собирательные трубочки почек. Клетки этих протоков относительно непроницаемы для воды, и в отсутствие АДГ моча не концентрируется и может выделяться в количествах, превышающих 20 л в сутки (норма 1—1,5 л в сутки). Для АДГ существуют два типа рецепторов – V1 и V2. Рецептор V2 обнаружен только на поверхности эпителиальных клеток почек. Связывание АДГ с V2 сопряжено с аденилатциклазной системой и стимулирует активацию протеинкиназы А (ПКА). ПКА фосфорилирует белки, которые стимулируют экспрессию гена мембранного белка — аквапорина-2. Аквапорин 2 перемещается к апикальной мембране, страивается в нее и образует водные каналы. Эти обеспечивают селективную проницаемость мембраны клеток для воды. Молекулы воды свободно диффундируют в клетки почечных канальцев, а затем поступают в интерстициальное пространство. В результате происходит реабсорбция воды из почечных канальцев. РецепторытипаV1 локализованы в мембранах гладких мышц. Взаимодействие АДГ с рецептором V1 приводит к акти-вации фосфолипазы С, которая гидролизует фосфатидилинозитол-4,5-бифоcфат с образованием ИФ-3. ИФ-3 вызывает высвобождение Са2+ из эндоплазматического ретикулума. Результатом действия гормона через рецепторы V1 является сокращение гладкомышечного слоя сосудов. Дефицит АДГ, вызванный дисфункцией задней доли гипофиза, а также нарушением в системе передачи гормонального сигнала может приводить к развитию несахарного диабета. Основным проявлением несахарного диабета является полиурия, т.е. выделение большого количества мочи низкой плотности. Альдостерон- наиболее активный минералокортикостероид синтезируется в коре надпочечников из холестерина. Синтез и секрецию альдостерона клетками клубочковой зоны стимулируют ангиотензин II, АКТГ, простагландин Е. Эти процессы также активируются при высокой концентрации К+ и низкой концентрации Na+. Гормон проникает внутрь клетки-мишени и взаимодействует со специфическим рецептором, расположенным как в цитозоле, так и в ядре. В клетках почечных канальцев альдостерон стимулирует синтез белков, которые выполняют разные функции. Эти белки могут: а) увеличивать активность натриевых каналов в мембране клеток дистальных почечных канальцев, способствуя тем самым транспорту ионов натрия из мочи в клетки; б)являться ферментами ЦТК и, следовательно, увеличивать способность цикла Кребса генерировать молекулы АТР, необходимые для активного транспорта ионов; в) активировать работу насоса К+, Na+-АТФазы и стимулировать синтез новых насосов. Суммарным результатом действия белков, которых индуцируется альдостероном, является увеличение реабсорбции ионов натрия в ка-нальцах нефронов, что вызывает задержку NaCl в организме. Главным механизмом регуляции синтеза и секреции альдостерона служит ренин-ангиотензиновая система. Ренин — фермент, продуцируемый юкстагломерулярными клетками почечных афферентных артериол. Локализация этих клеток делает их особенно чувствительными к изменению артериального давления. Снижение артериального давления, потеря жидкости или крови, уменьшение концентрации NaCI стимулируют высвобождение ренина.Ангиотензиноген —a2 - глобулин, образующийся в печени. Он служит субстратом для ренина. Ренин гидролизует пептидную связь в молекуле ангиотензиногена и отщепляет N-концевой декапептид (ангиотензин I).Ангиотензин I служит субстратом для антиотензинпревращающего фермента карбоксидипептидилпептидазы, выявленного в эндотелиальных клетках и плазме крови. От ангиотензина I отщепляются 2 терминальные аминокислоты с образованием октапептида — ангиотензина II.Ангиотензин II стимулирует выработку альдостерона, вызывает сужение артериол вследствие чего повышается артериальное давление и вызывает жажду. Ангиотензин II активирует синтез и секрецию альдостерона через инозитолфосфатную систему. ПНФ — пептид, содержащий 28 аминокислот с единственным дисульфидным мостиком. ПНФ синтезируется и хранится в виде препрогормона (состоящего из 126 аминокислотных остатков) в кардиоцитах.Основной фактор, регулирующий секрецию ПНФ — повышение артериального давления. Другие стимулы: увеличение осмомолярности плазмы, повышение частоты сердцебиения, повышенный уровень катехоламинов в крови и глюкокортикоидов. Основные органы-мишени ПНФ - почки, периферические артерии. Механизм действия ПНФ имеет ряд особенностей. Рецептор ПНФ плазматической мембраны являем белком, обладающим активностью гуанилатциклазы. Рецептор имеет доменное строение. Домен, связывающийся с лигандом, локализован во внеклеточном пространстве. В отсутствие ПНФ внутриклеточный домен рецептора ПНФ находится в фосфорилированном состоянии и неактивен. В результате связывания ПНФ с рецептором гуанилатциклазная активность рецептора возрастает и происходит образование циклического GMP из GTP. В результате действия ПНФ ингибируются образование и секреция ренина и альдостерона. Суммарным эффектом действия ПНФ является увеличение экскреции Na+ и воды и понижение артериального давления. ПНФ обычно рассматривают как физиологический антагонист ангиотензина II, поскольку под его влияния возникают не сужение просвета сосудов и (через регуляцию секреции альдостерона) задержка натрия, а наоборот, расширение сосудов и потеря соли.
Внешние и внутренние пути свертывания кровиВо внешнем пути свертывания крови участвуют тромбопластин (тканевой фактор, фактор III), проконвертин (фактор VII), фактор Стюарта (фактор X), проакцелерин (фактор V), а также Са2+ и фосфолипиды мембранных поверхностей, на которых образуется тромб. Гомогенаты многих тканей ускоряют свёртывание крови: это действие называют тромбопластиновой активностью. Вероятно, она связана с наличием в тканях какого-то специального белка. Факторы VII и X - проферменты. Они активируются путём частичного протеолиза, превращаясь в протеолитические ферменты - факторы VIIа и Xа соответственно. Фактор V – это белок, который при действии тромбина превращается в фактор V', который не является ферментом, но активирует фермент Xа по аллостерическому механизму; активация усиливается в присутствии фосфолипидов и Са2+. В плазме крови постоянно содержатся следовые количества фактора VIIа. При повреждении тканей и стенок сосуда освобождается фактор III – мощный активатор фактора VIIа; активность последнего увеличивается более чем в 15000 раз. Фактор VIIа отщепляет часть пептидной цепи фактора X, превращая его в фермент - фактор Xа. Сходным образом Xа активирует протромбин; образовавшийся тромбин катализирует превращение фибриногена в фибрин, а также превращение предшественника трансглутаминазы в активный фермент (фактор XIIIа). Этот каскад реакций имеет положительные обратные связи, усиливающие конечный результат. Фактор Xа и тромбин катализируют превращение неактивного фактора VII в фермент VIIа; тромбин превращает фактор V в фактор V', который вместе с фосфолипидами и Са2+ в 104–105 раз повышает активность фактора Xа. Благодаря положительным обратным связям скорость образования самого тромбина и, следовательно, превращения фибриногена в фибрин нарастают лавинообразно, и в течение 10-12 с кровь свёртывается. Свёртывание крови по внутреннему механизму происходит значительно медленнее и требует 10-15 мин. Этот механизм называют внутренним, потому что для него не требуется тромбопластин (тканевой фактор) и все необходимые факторы содержатся в крови. Внутренний механизм свёртывания также представляет собой каскад последовательных активаций проферментов. Начиная со стадии превращения фактора X в Xа, внешний и внутренний пути одинаковы. Как и внешний путь, внутренний путь свёртывания имеет положительные обратные связи: тромбин катализирует превращение предшественников V и VIII в активаторы V' и VIII', которые в конечном итоге увеличивают скорость образования самого тромбина. Внешний и внутренний механизмы свёртывания крови взаимодействуют между собой. Фактор VII, специфичный для внешнего пути свёртывания, может быть активирован фактором XIIа, который участвует во внутреннем пути свёртывания. Это превращает оба пути в единую систему свёртывания крови. Различают три механизма активации ферментов каскада:. Частичный протеолиз. Взаимодействие с белками-активаторами.Взаимодействие с клеточными мембранами. Ферменты прокоагулянтного пути содержат γ-карбоксиглутаминовую кислоту. Радикалы карбоксиглутаминовой кислоты образуют центры связывания ионов Са2+. В отсутствие ионов Са2+кровь не свертывается.
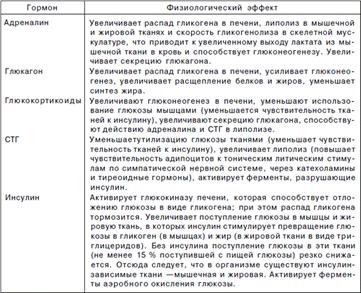
Гомеостаз. Гемостаз (свертывающая система крови) - это сложная система гомеостаза, которая с одной стороны поддер - живает кровь в жидком состоянии, обеспечивая нормальное кровоснабжение органов и тканей, а с другой стороны - останавливает кровотечение и предупреждает потерю крови из организма за счет поддержания структурной целостности стенок кровеносных сосудов и быстрого тромбообразования при их повреждении. Значение этой системы в том, что она препятствует потере крови из циркуляторного русла и тем самым обеспечивает нормальное кровоснабжение органов и тканей. Реализуется гемостаз 3-мя взаимодействующими между собой структурными компонентами: 1. Стенками кровеносных сосудов (в первую очередь их интимой (tunica intima)), 2. Клетками крови, 3-х плазменными ферментными системами (свертывающей, фибринолитической (плазминовой), калликреинкининовой и др.). Эндотелий представляет собой монослой клеток, непрерывно выделяющих на свою поверхность разнообразные биологически активные молекулы, обеспечивающие гемостаз. Среди этих молекул имеются как антикоагулянты, так и прокоагулянты. Для понимания роли эндотелия в гемостазе необходимо принять два его состояния – поврежденное (активное) и неповрежденное (спокойное). В неповрежденном (спокойном) состоянии эндотелий интактен, т.е. он не смачивается, его ламинарная сторона заряжена отрицательно, что отталкивает тромбоциты, он активно образует антикоагулянтные вещества. Тромбоциты – это клетки, ответственные за формирование первичной тромбоцитарной пробки в зоне повреждения за счет адгезии и последующей агрегации. Свои задачи тромбоциты выполняют благодаря разнообразным молекулам, секретируемым из внутриклеточных гранул, и при участии рецепторов к стимуляторам и ингибиторам коагуляции. Тромбоцитарные факторы свертывания – из них наиболее важен мембранный фосфолипидный фактор 3, который служит матрицей для взаимодействия плазменных факторов гемокоагуляции и образования их активных комплексов.По своим свойствам этот компонент идентичен кефалину и мембранному фактору эритроцитов - эритроцитину. Также важен 6-й фактор тромбоцитов - ретрактозим, необходимый для сокращения и уплотнения сгустка фибрина. В тромбоцитах имеются активаторы полимеризации мономеров фибрина, фактор V, а на поверхности и в их каналах концентрируются многие плазменные факторы свертывания и фибринолиза, и поэтому в гемостатической пробке создается их высокая концентрация (протромбин, тромбопластин, Ac-глобулин, конвертин, факторы II, III, V, VIII, IX, X, XI, XII, плазминоген и др. Поэтому тромбоциты наиболее существенно влияют на интенсивность и скорость локального свертывания в зоне тромбообразования, а не на процесс свертывания крови вообще. Плазменные факторы (многочисленные протеазы и их белковые коферменты) можно подразделить на три основные группы: белки системы свертывания, белки системы фибринолиза, белки калликреин-кининовой системы и системы комплемента. Активация каждой из этих систем является многоэтапным каскадным ферментативным процессом. Существуют общие особенности плазменных факторов гемостаза: 1. Практически все они находятся в крови в неактивной форме и только небольшое количество (около 1-2%) фактора VII циркулирует в активном виде. Активация белков гемостаза происходит механизмом частичного или ограниченного протеолиза. Ограниченный протеолиз способны осуществлять ферментативные факторы II, VII, IX, X, XI, XII, плазмин.2. Реакции свертывания не могут происходить непосредственно в кровотоке, им требуется отрицательно заряженная твердая поверхность – мембраны активированных тромбоцитов, фибробластов, макрофагов, бактерий и др. Такая поверхность носит название тромбопластин. Тромбопластин представляет собой комплекс белка гликопротеина (TF, тканевой фактор) и аминофосфолипида (фосфатидилсерина или фосфатидилэтаноламина), по происхождению выделяют тромбоцитарный и тканевой тромбопластин.3. Многие реакции также нуждаются в наличии ионов кальция, который обеспечивает связывание факторов свертывания с поверхностью мембран. В гепатоцитах происходит синтез факторов свертывания крови I, II, V, VII, VIII, IX, X, XI, XII, антикоагуляционных протеинов С и S, антитромбина III, плазминогена, прекалликреина, высокомолекулярного кининогена. Многие из этих соединений синтезируются также в клетках, непосредственно участвующих в гемостазе – тромбоцитах, эндотелии, лейкоцитах. Общей характерной особенностью факторов II, VII, IX, X, протеинов С и S является наличие в составе γ-карбоксиглутаминовой кислоты. γ-Карбоксиглутамат позволяет витамин-К-зависимым белкам при участии ионов Са2+ связываться с кислыми фосфолипидами на поверхности активированных клеток. Эта модифицированная аминокислота образуется при посттрансляционной модификации указанных белков ферментом γ-глутамилкарбоксилазой, ее коферментом является витамин К. Поступивший в печень витамин К (нафтохинон) восстанавливается НАДФH-зависимой витамин К-редуктазой с образованием дигидрохинона витамина К. Затем, в ходе ферментативной реакции, дигидрохинон окисляется и образуется неактивный 2,3-эпоксид витамина К. Регенерация эпоксида в дигидрохинон осуществляется редуктазами, коферментом которых является белок тиоредоксин. Образование фибрин полимера.После появления большого количества тромбина начинается превращение фибриногена в фибрин. При этом тромбин отщепляет конечные последовательности А и В в α- и β цепях фибриногена с образованием растворимых фибрин-мономеров. После этого спонтанно начинается латеральная ассоциация фибрин-мономеров с образованием олигомеров и, в конечном итоге, еще непрочной фибриновой сети (фибрин-полимер). Далее фактор XIIIa (фибрин-стабилизирующий фактор, трансглутаминаза) в присутствии ионов Са2+ образует амидные связи между остатками лизина одной молекулы фибрина-полимера и остатками глутамина другой молекулы, чем связывает нити фибрин-полимера друг с другом. Ковалентно сшитые между собой нити фибрина образуют прочную трехмерную сеть, в которую включены тромбоциты, эритроциты и лейкоциты. Также трансглутаминаза прикрепляет фибрин к фибронектину, прочно связанному с другими молекулами внеклеточного матрикса. В результате тромб оказывается прикрепленным к матриксу в области поврежденной стенки сосуда. В это время активированные тромбоциты, связанные с нитями фибрина через рецепторы GPIIb/IIIa, сокращаются под действием тромбостенина – происходит ретракция. Сгусток крови уплотняется, из него выдавливается часть сыворотки. Формирование окончательного тромба наступает на 10-15-й минуте после полимеризации фибрина. Нервная регуляцияконцентрации глюкозы в крови выражается в положительном влиянииn.vagus на секрецию инсулина и тормозящем влиянии на этот процесс симпатической иннервации. Кроме этого, выделение адреналина в кровь подвержено симпатическим влияниям. Основными факторами гормональнойрегуляции являютсяглюкагон, адреналин, глюкокортикоиды, соматотропный гормонс одной стороны, и инсулинс другой. Инсу- лин является единственным гормоном организма, действие которого нацелено на снижение уровня глюкозы крови. При его влиянии глюкозу поглощают мышцы и жировая ткань. Все остальные гормоны увеличивают гликемию, влияя на печень. Снижение концентрации глюкозы крови инсулиномдостигается следующими путями:
oувеличение синтеза гликогена– активация гликогенсинтазы и стимуляция ее синтеза, что облегчает превращение излишков глюкозы в гликоген, oактивация пентозофосфатного пути– индукция синтезаглюкозо-6-фосфат- дегидрогеназы и 6-фосфоглюконатдегидрогеназы, o усиление липогенеза– вовлечение глюкозы в синтез триацилглицеролов (см "Липиды", "Синтез триацилглицеролов"). Многие ткани совершенно нечувствительны к действию инсулина, их называют инсулиннезависимыми. К ним относятся нервная ткань, стекловидное тело, хрусталик, сетчатка, клубочковые клетки почек, эндотелиоциты, семенники и эритроциты. Глюкагонповышает содержание глюкозы крови: o увеличивая мобилизацию гликогеначерез активацию гликогенфосфорилазы, oстимулируя глюконеогенез– повышение работы ферментов пируваткарбоксилазы,фосфоенолпируват-карбоксикиназы,фруктозо-1,6-дифосфатазы. Адреналинвызывает гипергликемию: o активируя мобилизацию гликогена– стимуляция гликогенфосфорилазы, Глюкокортикоидыповышают глюкозу кровиoза счет подавления перехода глюкозы в клетку, oстимулируя глюконеогенез– увеличивают синтез ферментов пируваткарбоксилазы,фосфоенолпируват-карбоксикиназы,фруктозо-1,6-дифосфатазы.
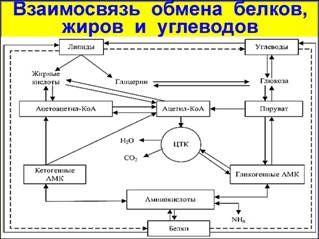
Взаимосвязь обменов углеводов, белков и жиров Обмен веществ в организме характеризуется тесной взаимосвязью между углеводами, белками и жирами. В организме белки, жиры и углеводы пищи в результате различных превращений теряют свои специфические свойства и часто образуют химические вещества одинаковой структуры. Углеводы, белки и жиры в пищеварительном тракте расщепляются до мономеров, которыми являются: моносахариды, аминокислоты, глицерин и жирные кислоты. В клетках организма происходит их дальнейший распад с образованием ряда конечных продуктов, в том числе CH3CO~SKoA. Последний образуется при окислении глюкозы, распаде жирных кислот, дезаминировании аминокислот. С другой стороны, ацетил-Ко А независимо от пути его образования участвует в синтезе определенных для каждого организма веществ: жирных кислот, углеводов, холестерина, желчных кислот, некоторых гормонов, витамина D. Он также служит одним из основных поставщиков энергии, главным "горючим" организма, так как при окислении его в цикле Кребса выделяется большое количество энергии, которая в виде АТФ обеспечивает жизнедеятельность всех клеток нашего тела. Поэтому, например, при недостатке в пище жиров, дефицит СН3СО ~ SKoA будет покрываться за счет повышенного распада углеводов и белков. Таким образом, нарушение любого процесса, приводящего к образованию ацетил-КоА, отразится на деятельности других видов обменов веществ, а следовательно, и организма в целом, а в ряде случаев может привести и к развитию патологии, как в случае сахарного диабета. Это - общий путь, характеризующий взаимосвязь обменов углеводов, белков и жиров в организме. Однако существуют и другие биохимические механизмы взаимосвязи между этими веществами.
Сущность дыхательной функции крови состоит в доставке кислорода от легких к тканям и углекислого газа от тканей к легким. Кровь осуществляет дыхательную функцию прежде всего благодаря наличию в ней гемоглобина. Физиологическая функция гемоглобина как переносчика кислорода основана на способности обратимо связывать кислород. Поэтому в легочных капиллярах происходит насыщение крови кислородом, а в тканевых капиллярах, где парциальное давлениекислорода резко снижено, осуществляется отдача кислорода тканям. В состоянии покоя ткани и органы человека потребляют около 200 мл кислорода в минуту. При тяжелой физической работе количество потребляемого тканями кислорода возрастает в 10 раз и более (до 2–3 л/мин). Доставка от легких к тканям такого количества кислорода в виде газа, физически растворенного в плазме, невозможна вследствие малойрастворимости кислорода в воде и плазме крови. Итак, функцию переносчика кислорода в организме выполняет гемоглобин. Строение гемоглобина Гемоглобин представляет собой белок, включающий 4 гемсодержащие белковые субъединицы. Между собой протомеры соединяются гидрофобными, ионными, водородными связями по принципу комплементарности.Механизм образования оксигемоглобина. При взаимодействии молекулы кислорода с одним из четырех гемов гемоглобина кислород присоединяется к одной из половинок молекулы гемоглобина (допустим, к α-цепи этой половинки). Как только такое присоединение произойдет, α-полипептидная цепь претерпевает конформа-ционные изменения, которые передаются на тесно связанную с ней β-цепь; последняя также подвергается конформационным сдвигам. β-Цепь присоединяет кислород, имея уже большее сродство к нему. Таким путем связывание одной молекулы кислорода благоприятствует связыванию второй молекулы (так называемое кооперативное взаимодействие). После насыщения кислородом одной половины молекулы гемоглобина возникает новое, внутреннее, напряженное состояние молекулы гемоглобина, которое вынуждает и вторую половину гемоглобина изменить конфор-мацию. Теперь еще две молекулы кислорода, по-видимому, по очереди связываются со второй половинкой молекулыгемоглобина, образуя оксигемоглобин. Роль 2,3-дифосфоглицерата 2,3-Дифосфоглицерат образуется в эритроцитах из 1,3-дифосфоглицерата, промежуточного метаболита гликолиза, в реакциях, получивших название шунт Раппопорта. 2,3-Дифосфоглицерат располагается в центральной полости тетрамера дезоксигемоглобина и связывается с β-цепями, образуя поперечный солевой мостик между атомами кислорода 2,3-дифосфоглицератаи аминогруппами концевого валина обеих β-цепей, также аминогруппами радикалов лизинаи гистидина. Функция 2,3-дифосфоглицерата заключается в снижении сродства гемоглобина к кислороду. Это имеет особенное значение при подъеме на высоту, при нехватке кислорода во вдыхаемом воздухе. В этих условиях связывание кислорода с гемоглобином в легких не нарушается, так как концентрация его относительно высока. Однако в тканях за счет 2,3-дифосфоглицерата отдача кислорода возрастает в 2 раза.
Ферменты плазмы крови По происхождению ферменты плазмы крови можно подразделить на 3 группы. 1. Собственные ферменты плазмы крови (секреторные). Они образуются в печени, но проявляют своё действие в крови. К ним относятся ферменты свертывающей системы крови – протромбин, проакцелерин, проконвертин, а также церулоплазмин, холинэстераза. 2. Экскреторные ферменты – попадают в кровь из различных секретов – дуоденального сока, слюны и т.д. К ним относятся амилаза, липаза. 3. Клеточные ферменты – попадают в кровь при повреждениях или разрушениях клеток или тканей.
Гормоны половых желез Гонады - яйчники и семенники - железы смешанного типа. По химической природе их гормоны - стероиды. Андрогены (мужские) синтезируются в семенниках и коре надпочечников. Эстрогены (женские) - в яичниках и коре надпочечников. Синтезируются из холестерола, который образуется из Ац-КоА. Ац-КоА® холестерол® прегненолон ®прогестерон ®кортикостероиды. Из прогестерона также образуются андрогены (тестостерон) , а из них - эстрогены. Эстрогены: эстрадиол, эстрон (образуется из эстрадиола в плаценте), эстриол, прогестерон.
Андрогены: тестостерон, андростерон (образуется из тестостерона).
Органы-мишени для эстрадиолов - матка и молочные железы, для андрогенов - простата, семенные пузырьки, мышцы. Механизм действия – цитозольный - гормоны соединяются с цитозольными рецепторами и действуют на ген. Основной эффект - синтез специальных белков репродуктивной системы.
Влияние на обмен веществ. Эстрогены: 1. синтезируют специфические белки; 2. обуславливают положительный азотистый баланс; 3. активируют пентозный цикл; 4. препятствуют накоплению липидов в жировой ткани и печени. Эстрогены имеют цикличность секреции: в фолликулярной фазе синтезируются эстрогены, в лютеиновой фазе - прогестерон.
Андрогены: 1. синтезируют специальные белки половой сферы; 2. стимулируют синтез мышечных белков; 3. стимулируют синтез белков печени и почек; 4. ускоряют развитие костной ткани; 5. ускоряют аэробное окисление; 6. обладают анаболическим эффектом;
Гормоны коркового вещества надпочечников (кортикостероиды). Известно более 30 гормонов-стероидов, т.е. производные циклопентанпергидрофенантрена: 1. глюкокортикоиды - оказывают влияние на углеводный обмен; 2. минералокортикоиды - на минеральный обмен; 3. половые гормоны. Глюкокортикоиды: кортикостерон, кортизол (самый активный в организме человека), кортизон.
Клетки-мишени для глюкокортикоидов - печень, почки, лимфоидная ткань, соединительная ткань, мышцы. Рецепторы находятся в цитозоле, проходят через мембрану и действуют на ген. Гормон ® ген ® белок. Влияние на обмен веществ: 1. активация глюконеогенеза; 2. повышение уровня глюкозы в крови; 3. повышение синтеза гликогена в печени; 4. стимулируют липолиз в области конечностей и липогенез в области туловища и лица; 5. повышение окисления жирных кислот; 6. повышение образования кетоновых тел; 7. в печени увеличивается синтез белка, в мышцах, лимфоидной, соединительной тканях увеличивается распад белка; 8. противовоспалительное действие, вызывают инволюцию лимфоидной ткани; 9 антиаллергический эффект, подавляют образование антител; 10. подавление синтеза белка в соединительной ткани, задержка образования рубцов и спаек. Использование глюкокортикоидов в клинике - противовоспалительные, при трансплантации органов для снижения образования рубцов и спаек. Глюкокортикоиды влияют и на минеральный обмен, но в меньшей степени.
Минералокортикоиды: - диоксикортикостерон; - альдостерон.
Клетки-мишени - дистальные канальцы почек. Рецепторы находятся внутри клеток – цитозольные рецепторы. Влияют на синтез белков, транспорт натрия через мембраны, т.е. повышают реабсорбцию натрия и хлоридов из первичной мочи, задерживая натрий в организме. Влияют на углеводный обмен, как и гипоталамус, но в меньшей степени. Гипофункция коры надпочечников - болезнь Адиссона (бронзовая болезнь). Снижается устойчивость организма к стрессам, гипогликемия, потеря натрия и накопление калия, гипотония, мышечная слабость, утомляемость, повышение пигментации кожи, возможна гибель из-за нарушения водно-солевого обмена. Гиперфункция - синдром Иценко-Кушинга
Гормоны коры и мозгового вещества надпочечников Гормоны мозгового слоя- адреналин и норадреналин - производные тирозина. [схема образования: тирозин ® (оксигеназа, 1/2О2)диоксифенилаланин ® (декарбоксилаза, -СО2)дофамин ® (+О2)норадреналин ® (+СН3)адреналин].
Органы-мишени - печень, скелетные мышцы, сердечная мышца, слюнные железы, матка. Механизм: через аденилатциклазную систему. Увеличивается цАМФ ® увеличение протеинкиназы ® увеличение фосфорилирования. Фосфорилазы действуют на превращение гликогена в глюкозу. Адреналин действует и в печени, и в мышцах, вследствие чего повышается содержание глюкозы в крови, повышается содержания молочной кислоты. Адреналин повышает потребление кислорода, увеличивает липолиз, что приводит к росту количества свободных жирных кислот в крови. Повышает кровяное давление, частоту сердечных сокращений. Адреналин действует на a1, a2, b1 и b2-адренорецепторы. Если гормоны связаны с b-рецепторами, то происходит активация цАМФ, если с a-рецепторами - ингибирование цАМФ. Классификация гормонов Существует несколько видов классификации. По месту образования гормонов: 1. гормоны гипоталамуса; 2. гормоны гипофиза; 3. гормоны щитовидной железы; 4. гормоны поджелудочной железы; 5. гормоны паращитовидных желез; 6. гормоны надпочечников; 7. гормоны половых желез; 8. гормоны местного действия. По химическому строению: 1. белково-пептидные гормоны: гормоны гипоталамуса, гипофиза, поджелудочной железы, паращитовидных желез; 2. производные аминокислот: адреналин, норадреналин, тироксин, трийодтиронин; 3. стероиды: в их основе лежит структура циклопентанпергидрофенантрена, образуются из холестерина (половые гормоны, коры надпочечников). По механизму действия (по расположению рецепторов): 1. гормоны, действующие через внутриклеточный рецептор - липофильные гормоны - стероиды и тиреоидные гормоны; 2. гормоны, действующие через рецепторы, находящиеся на поверхности клетки - гидрофильные гормоны. Они действуют через внутриклеточный посредник - мессенджер. Гормон - первый посредник, а цАМФ, ионы Са2+, фосфатидилинозиды - вторые (чаще цАМФ, которая образуется из АДФ) посредники. [рис. цАМФ] Сахарный диабет Выделяют: 1. Инсулин - зависимый сахарный диабет - абсолютная недостаточность инсулина - 1 тип. При диабете первого типа снижается или перестаёт вырабатываться инсулин поджелудочной железой. В основном, причиной появления диабета 1 типа становится генетическая предрасположенность. При этом генетически человеку передаётся не сам диабет, а предрасположенность к нему. 2. Инсулин - независимый сахарный диабет. Протекает с относительной недостаточностью инсулина. В крови у таких больных инсулин в норме или повышен. Может быть с ожирением и с нормальной массой тела. У такого сахарного диабета причиной возникновения считается снижение количества рецепторов к инсулину, а также недостаток внутриклеточных ферментов, приводящий к нарушению обмена глюкозы в клетках и тканях организма. Устойчивость периферических тканей к гормону поджелудочной железы — инсулину приводит к инсулинизму (повышению секреции инсулина), что также способствует снижению числа рецепторов и появлению симптомов сахарного диабета у женщин и мужчин.1) затруднение транспорта глюкозы в мышечную и жировую ткань; 2) угнетение окисления глюкозы по пути фосфорилирования в связи со снижением активности ключевых ферментов, превращения глюкозы (гексокиназы, гликокиназы);3) понижение синтеза гликогена в печени в связи со снижением активности гликокиназы;4) усиление глюконеогенеза. Следствием всех этих прцессов является развитие основного симптома сахарного диабета – гипергликемии. Основной путь превращения глюкозы в физиологических условиях – это путь окислительного фосфорилирования, осуществляемый под действием инсулина. В условиях его дефицита процесс окисления глюкозы по пути фосфорилирования угнетается и увеличивается удельный вес других путей обмена глюкозы. В частности, начинает преобладать анаэробное расщепление глюкозы. В результате в тканях образуется в повышенном количестве молочная кислота. Выделение ее в кровь ведет к развитию гиперлактацедемии, усугубляющей диабетический ацидоз. Кроме того, усиливается превращение глюкозы по сорбитоловому пути и, соответственно, накапливаются продукты этого превращения (сорбитол и фруктоза). Сорбитоловый путь превращения глюкозы характерен для хрусталика глаза, нервной ткани, эндотелия сосудов. Накопление сорбитола и фруктозы в тканях способствует развитию осложнений сахарного диабета (катаракта, полинейропатия, ангиопатия). При диабете увеличивается также использование глюкозы в образовании гликопротеидов (белков, составляющих базальную мембрану сосудов), что играет важную роль в патогенезе микроангиопатий. Гипергликемия вызывает гликозилирование различных белков: гемоглобин, альбумин, белки базальной мембраны сосудов, что приводит к изменению их свойств, повышению иммуногенности и имеет значение в развитии сосудистых поражений.Повышение уровня глюкозы в крови выше почечного порога (9,5-10 ммоль/л) сопровождается выделением сахара с мочой – глюкозурией, которая тем выше, чем интенсивнее гипергликемия. Выделение глюкозы с мочой сопроваждается увеличением диуреза. Глюкоза увлекает за собой жидкость в связи с повышением осмотического давления в провизорной моче и снижением реабсорбции мочи в канальцах почек. На каждый грамм глюкозы выделяется 20-40 мл жидкости. Таков механизм полиурии. Полидипсия при диабете вторичного происхождения. Она связана с интенсивным обезвоживанием организма. Одновременно нарушается ВОДНО-СОЛЕВОЙ ОБМЕН. Больной теряет калий и натрий, усиление диуреза приводит к дегидратации. НАРУШЕНИЕ ЖИРОВОГО ОБМЕНА при дефиците инсулина сводится к снижению синтеза жира и усилению липолиза. В результате мобилизации жира из депо развивается гиперлипидемия. Избыточный жир откладывается в бедных гликогеном гепатоцитах, вызывая жировую инфильтрацию печени. В кровь выделяются в повышенных количествах неэстерифицированные жирные кислоты, заменяющие глюкозу в качестве энергетического материала.В печени, в условиях пониженного содержания гликогена, уменьшено превращение ацетил-Кол, а в цикле Кребса образуются в повышенном количестве недоокисленные продукты жирового обмена – кетоновые тела (оксимасляная кислота, ацетоуксусная кислота, ацетон). Развивается характерный для декомпенсации сахарного диабета кетоацидоз. НАРУШЕНИЕ БЕЛКОВОГО ОБМЕНА заключается в угнетении синтеза белка и повышенном его распаде. Вследствие угнетения синтеза белка в детском возрасте наблюдается задержка роста.В печени белок интенсивно превращается в углеводы. В процессе этого превращения диспротеинемии в сторону преобладания глобулиновых фракций образуются продукты распада (аммиак, мочевина, аминокислоты). Поступая в кровь, они обуславливают гиперазотемию.При сахарном диабете вследствие нарушения белкового обмена снижается продукция защитных белков, что приводит к снижению иммунитета. Компенсация нарушений углеводного обмена, как правило, значительно улучшает показатели белкового, жирового и водно-солевого обмена. Однако нередко в клинической практике приходится прибегать к специальному лечению этих нарушений.Нарушение функции белков (гемоглобина, эндотелия сосудов, почечного эпителия), анемия, гипоальбунемия, нарушение микроциркуляции и трофики тканей.
Белки иммунной системы В иммунном ответе можно выделить два основных процесса: 1)распознавание чужеродных молекул, т. е. различение их от собственных макромолекул организма; 2) обезвреживание (разрушение) чужеродных молекул, паразитарных организ мов, мутантных клеток. Ядро иммунной системы составляют три семейства белков: 1)иммуноглобулины (антитела), синтезируемые и секретируемые в кровь лим фоцитами В; 2)l'-рецепторы,которые синтезируются лимфоцитами T и локализуются на наружной поверхности их плазматической мембраны;3)белки главного комплекса гистосовместимости (белки ГКГ), синтезируемые многими (почти всеми) клетками организма и локализованные тоже на по верхности клеток.СТРОЕНИЕ АНТИТЕЛ Антитела или иммуноглобулины (Ig) синтезируются в лимфоцитах В, главным об разом в лимфатических узлах и селезенке, и выделяются в кровь, образуя фракцию иммуноглобулинов плазмы. Структурную основу иммуноглобулинов составляют четыре пептидные цепи, соединенные друг с другом дисульфидными связями: две тяжелые (цепи Н) с мо лекулярной массой 50 ООО (от 450 до 700 аминокислотных остатков) и две легкие (цепи L) с молекулярной массой 25 000 (около 200 аминокислотных остатков) (рис. 20.1). Такую структуру обычно называют мономером. В пептидных цепях различают вариабельные (V) и постоянные, или константные (С), области. По различиям первичной структуры постоянных областей легкие цепи делятся на два типа (к и ^), тяжелые — на пять типов (табл. 20.1). По типу тяжелых цепей, входящих в мономеры, все иммуноглобулины делятся на пять классов. Каждый класс включает огромное множество индивидуальных иммуноглобулинов, разли чающихся по первичной структуре вариабельных областей; общее число инди видуальных иммуноглобулинов всех классов равно примерно IO7. Молекулы ан тител классов IgG, IgD и IgE мономерны; они имеют Y-образнуюформу. Антите ла IgA построены издвух-четырехмономеров (H,L2)24), a IgM — из пяти мономеров (H2L2)rj. Гормоны передней доли гипофиза (аденогипофиза): 1. простые белки; 2. гликопротеины; 3. пептиды семейства проопиомеланокортина (ПОМК).
Гормоны - простые белки: 1. гормон роста (ГР); 2. пролактин (ПРЛ); 3. плацентарный пролактин (лактоген) (ПЛ) - хорионический соматомаммотропин (ХС). Эти гормоны стимулируют рост и лактацию. Содержат 190-195 аминокислот, они близки по строению и эффекту. Гормон роста содержит 191 АК. Строение на 85% аналогично строению холестерина (ХС) и на 35%- с пролактином (ПРЛ). Концентрация гормона роста выше, чем других гормонов. Секретируется эпизодически. Влияют стрессовые состояния, белковая пища, физические упражнения – при этом повышается секреция. Влияние на обмен веществ: а) повышает синтез белка: - повышает транспорт АК в клетки; - повышает синтез РНК и ДНК; - повышает интенсивность трансляции белков. Гормон роста действует через инсулинподобный фактор роста (ИФР-1,2). ИФР-1- это главный посредник в действии гормона роста. б) влияет на углеводный обмен - антагонист инсулина: - повышает содержание глюкозы в крови - гипергликемия; - снижает периферическую утилизацию; - повышает продукцию глюкозы в процессе глюконеогенеза. в) влияет на липидный обмен, стимулирует липолиз: - увеличивает скорость окисления ВЖК (в печени). г) влияет на минеральный обмен: - задерживает в организме кальций, магний, фосфор, стимулирует рост костной ткани. Гиперсекреция гормона роста у детей проявляется в виде гигантизма, у взрослых - акромегалия. Дефицит - гипосекреция - приводит к карликовости.
Пролактин - белок, состоящий из 198 АК. Его секреция увеличивается при беременности и лактации. Участвует в инициации лактации и поддерживает ее уровень. Внутриклеточный медиатор для ПЛ не известен.
Хорионический соматомаммолиберин (плацентарный лактоген) - не выполняет строго специфических функций, стимулирует лактацию и лютеотропную активность. Эффекты метаболизма такие же, как и у гормона роста. Семейство проопиомеланокортина (ПОМК) ПОМК - это молекула-предшественник, состоит из 285 АК-остатков. Этот белок гидролизуется до пептидов, которые и имеют гормональный эффект. Выделяют 3 группы пептидов: 1. АКТГ (адренокортикотропный гормон) Из него могут образовываться a-МСГ (меланоцитстимулирующий гормон) и кортикотропиноподобный пептид; 2. b-липотропин - из него образуются a-липотропин, b-МСГ, b-эндорфины (из него - a-эндорфины, g-эндорфины); 3. N-концевой пептид, из него образуется g-МСГ.
АКТГ - полипептид, состоит из 39 АК. Клетки-мишени - клетки коры надпочечников. Повышает синтез стероидов коры надпочечников за счет превращения холестерола в прегненалон. Его содержание значительно возрастает - это предшественник кортикостероидов. Повышает синтез белков и РНК в коре надпочечников. Внутриклеточный посредник - цАМФ. АКТГ стимулирует синтез аденилатциклазы в надпочечниках и жировой ткани, следовательно, интенсивность липолиза повышается. При избытке АКТГ - синдром Кушинга - повышается пигментация кожи, наблюдается отрицательный азотистый баланс, отрицательный баланс К и Р, но задерживается в организме натрий, что приводит к развитию отеков, повышению давления. Гиперсекреция АКТГ повышает уровень глюкозы в крови и может развиться стероидный диабет, повышается уровень свободных жирных кислот в крови, так как повышается липолиз - это характерно для стрессовых ситуаций.
b-липотропин - стимулирует липолиз.
b-эндорфины - в гипофизе они ацилированны и неактивны, а в ЦНС они свободны и проявляют активность. Они выполняют нейромедиаторную функцию и нейрорегуляторную. Эндорфины имеют те же рецепторы, что и морфины. Эндорфины регулируют чувство удовольствия (счастья). Применение различных опиатов приводит к увеличению числа рецепторов и возникает зависимость от внешних веществ (если количество рецепторов снижается).
МСГ - стимулирует меланогенез, т.е. образование меланоцитов и меланина.
Гормоны задней доли гипофиза (нейрогипофиза): - вазопрессин (антидиуретический гормон); - окситоцин. Синтезируются в гипоталамусе, а выделяются из задней доли гипофиза. Вазопрессин - по структуре пептид, состоящий из 9 АК. Эти гормоны влияют на сокращения гладкой мышечной ткани, но клетки-мишени различны. Для вазопрессина это клетки сосудов и дистальных извитых канальцев и извитых трубочек почек. В связи с этим вазопрессин повышает обратное всасывание воды (реабсорбцию) в почках, т.е. диурез снижается и повышается сокращение гладкомышечных клеток сосудов. Он действует через вторичный посредник - цАМФ. При нарушении секреции вазопрессина может развиться несахарный диабет (несахарное мочеизнурение - нарушается реабсорбция и объем воды повышается - полиурия, но в моче нет глюкозы).
Окситоцин действует на гладкомышечную ткань матки и молочных желез. Посредник неизвестен. Используется для стимуляции родовой деятельности, оптимизирует секрецию молока.
Гормоны щитовидной железы В щитовидной железе образуются 3 гормона, 2 из которых близки по действию и образуются из тирозина: 1. 3,5,3’-трийодтиронин (Т3); 2. 3,5,3’,5’–тетрайодтиронин (Т4); 3. тиреокальцитонин. Т3 и Т4 образуются из АК – тирозина.
Образование тиреотропных гормонов происходит на белке тиреоглобулине. Сначала молекулы тирозина включаются в белковую структуру.
Особенности образования тиреоидных гормонов: - они связаны со структурой тиреоглобулина в ЩЖ; - для синтеза необходим йод. ЩЖ способна концентрировать йод в виде йодидов, при этом происходит: 1. окисление йодидов с помощью тиреопероксидазы; 2. йодирование тирозина; 3. конденсация (соединение) йод-тирозина. Тиреоглобулин, который содержит Т3 и Т4, разрушается с образованием этих гормонов под влиянием тиреотропного гормона (тиреоглобулин«(над стрелкой ТТГ, под - KI) Т3+Т4. KI тормозит распад тиреоглобулина и поэтому применяется для лечения гипертериоза. В кровь поступают свободные тиронины, но в крови они транспортируются с тироксин-связывающим глобулином (ТСГ). Биологически более активны свободные тиронины, но их немного.
Механизм действия тиреоидных гормонов. Тиреоидные гормоны проникают через плазматическую мембрану и связываются с ядерными рецепторами (рецепторы есть в ядре и цитоплазме). Сродство Т3 в 10 раз больше, чем Т4. Эти гормоны (Т3, Т4) индуцируют синтез белков, активируют транскрипцию мРНК.
Влияние на обмен веществ: 1. увеличивают поглощения кислорода почти во всех тканях, кроме мозга и половых желез; 2. повышают активность Na+,K+-АТФ-азы, но если активность повышается, концентрация АТФ снижается, следовательно усиливаются окислительные процессы; 3. снижают отношение кислорода к АТФ. Тиреоидные гормоны повышают синтез белка, при их нормальной концентрации наблюдается положительный азотистый баланс, но при очень высокой концентрации этих гормонов проявляется обратный эффект.
Нарушения секреции. Гипофункция – гипотиреоз - у детей может привести к нарушению умственного и физического развития, кретинизм. У взрослых - микседема (слизистый отек). Развитие эндемичного зоба связано с недостатком йодидов в воде и пище. Железистая ткань не развивается, а вместо нее образуется соединительная ткань, что приводит к увеличению массы ЩЖ. Гиперсекреция тиреоидных гормонов – гипертиреоз – проявляется в виде тиреотоксикоза - увеличение ЩЖ, но за счет железистой ткани. При этом повышаются окислительные процессы в организме. Наблюдается отрицательный азотистый баланс, потеря массы тела, повышается возбудимость, пучеглазие, дрожание конечностей.
Кальцитонин образуется в клетках ЩЖ. Это пептид, состоящий из 32 АК. Он снижают уровень Са2+ в крови, но механизм действия недостаточно изучен.
Гормоны паращитовидных желез Парат-гормон, состоит из 84 АК, регулирует уровень Са2+, стимулирует выход кальция (и фосфора) из костей в кровь; Повышают реабсорбцию кальция в почках, но стимулируется выход фосфора; Стимулирует образование кальцитриола из витамина Д3 в почках; Повышает всасывание кальция в кишечнике. Действует через цАМФ.
|
|||||||||||||||
|
|
Последнее изменение этой страницы: 2018-05-10; просмотров: 512. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |