|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
Лизосомы и комплекс ГольджиЛизосомы (гр. lysis – «разложение, распад» и soma – «тело») – это пузырьки диаметром 200–400 мкм. Имеют одномембранную оболочку, которая снаружи иногда бывает покрыта волокнистым белковым слоем. Содержат набор ферментов, которые осуществляют при низких значениях рН гидролитическое расщепление веществ (нуклеиновых кислот, белков, жиров, углеводов). Основная функция – внутриклеточное переваривание различных химических соединений и клеточных структур. Выделяют первичные (неактивные) и вторичные лизосомы (в них протекает процесс переваривания). Вторичные лизосомы образуются из первичных. Они подразделяются на гетеролизосомы и аутолизосомы.
Схема строения и микрофотография лизосом В гетеролизосомах (фаголизосомах) протекает процесс переваривания материала, который поступает в клетку извне путем активного транспорта (пиноцитоза и фагоцитоза).
В аутолизосомах (цитолизосомах) подвергаются разрушению собственные клеточные структуры, которые завершили свою жизнь. Вторичные лизосомы, которые уже перестали переваривать материал, называются остаточными тельцами. В них нет гидролаз, содержится непереваренный материал. При нарушении целостности мембраны лизосом или при заболевании клетки гидролазы поступают внутрь клетки из лизосом и осуществляют ее самопереваривание (автолиз). Этот же процесс лежит в основе процесса естественной гибели всех клеток (апоптоза). Комплекс Гольджи Пластинчатый комплекс Гольджи – это упаковочный центр клетки. Представляет собой совокупность диктиосом (от нескольких десятков до сотен и тысяч на одну клетку). Диктиосома – стопка из 3—12 уплощенных цистерн овальной формы, по краям которых расположены мелкие пузырьки (везикулы). Более крупные расширения цистерн дают вакуоли, содержащие резерв воды в клетке и отвечающие за поддержание тургора. Пластинчатый комплекс дает начало секреторным вакуолям, в которых содержатся вещества, предназначенные для вывода из клетки. При этом просекрет, поступающий в вакуоль из зоны синтеза, (ЭПС, митохондрии, рибосомы), подвергается здесь некоторым химическим превращениям. 
Строение и микрофотография комплекса Гольджи Комплекс Гольджи дает начало первичным лизосомам. В диктиосомах также синтезируются полисахариды, гликопротеиды и гликолипиды, которые затем идут на построение цитоплазматических мембран.
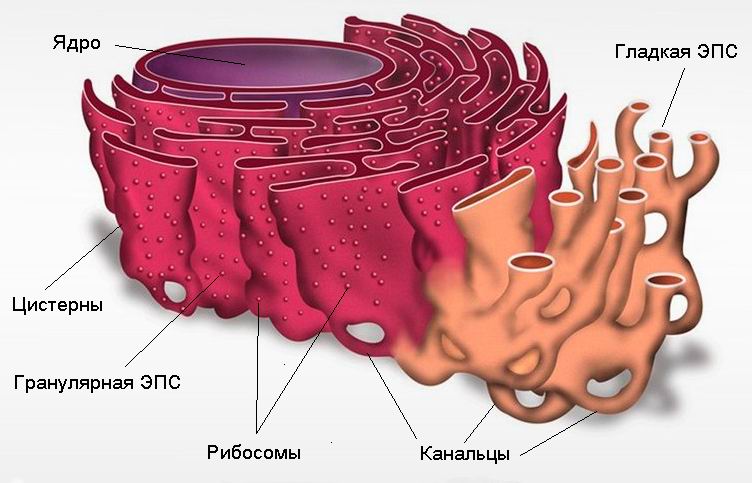
ЭНДОПЛАЗМАТИЧЕСКАЯ СЕТЬ Эндоплазматический ретикулум (ЭПС) – система сообщающихся или отдельных трубчатых каналов и уплощенных цистерн, расположенных по всей цитоплазме клетки. Они отграничены мембранами (мембранными органеллами). Иногда цистерны имеют расширения в виде пузырьков. Каналы ЭПС могут соединяться с поверхностной или ядерной мембранами, контактировать с комплексом Гольджи.
Схема строения и микрофотография эндоплазматической сети (ЭПС)
В данной системе можно выделить гладкую и шероховатую (гранулярную) ЭПС. Шероховатая ЭПС На каналах шероховатой ЭПС в виде полисом расположены рибосомы. Здесь протекает синтез белков, преимущественно продуцируемых клеткой на экспорт (удаление из клетки), например, секретов железистых клеток. Здесь же происходят образование липи-дов и белков цитоплазматической мембраны и их сборка. Плотно упакованные цистерны и каналы гранулярной ЭПС образуют слоистую структуру, где наиболее активно протекает синтез белка. Это место называется эргастоплазмой. Гладкая ЭПС На мембранах гладкой ЭПС рибосом нет. Здесь протекает в основном синтез жиров и подобных им веществ (например, стероидных гормонов), а также углеводов. По каналам гладкой ЭПС также происходит перемещение готового материала к месту его упаковки в гранулы (в зону комплекса Гольджи). В печеночных клетках гладкая ЭПС принимает участие в разрушении и обезвреживании ряда токсичных и лекарственных веществ (например, барбитуратов). В поперечнополосатой мускулатуре канальцы и цистерны гладкой ЭПС депонируют ионы кальция.
Немембранные органоиды В эту группу органоидов входят рибосомы, микротрубочки и микрофиламенты, клеточный центр. Рибосома Это округлая рибонуклеопротеиновая частица. Диаметр ее составляет 20–30 нм. Состоит рибосома из большой и малой субъединиц, которые объединяются в присутствии нити м-РНК (матричной, или информационной РНК). Комплекс из группы рибосом, объединенных одной молекулой м-РНК наподобие нитки бус, называется полисомой.
Схема строения и микрофотография рибосом (на фото – полисома) Эти структуры либо свободно расположены в цитоплазме, либо прикреплены к мембранам гранулярной ЭПС (в обоих случаях на них активно протекает синтез белка). Полисомы гранулярной ЭПС образуют белки, выводимые из клетки и используемые для нужд всего организма (например, пищеварительные ферменты, белки женского грудного молока). Кроме этого, рибосомы присутствуют на внутренней поверхности мембран митохондрий, где также принимают активное участие в синтезе белковых молекул. Клеточный центр Клетки всех животных, некоторых грибов, водорослей, низших растений характеризуются наличием клеточного центра. Клеточный центр обычно располагается рядом с ядром.
Схема строения и микрофотография клеточного центра Он состоит из двух центриолей, каждая из которых представляет собой полый цилиндр диаметром около 150 нм, длиной 300–500 нм. Центриоли расположены взаимно перпендикулярно. Стенка каждой центриоли образована 27 микротрубочками, состоящими из белка тубулина. Микротрубочки сгруппированы в 9 триплетов. Из центриолей клеточного центра во время деления клетки образуются нити веретена деления. Центриоли поляризуют процесс деления клетки, чем достигается равномерное расхождение сестринских хромосом (хроматид) в анафазе митоза. Микротрубочки Это трубчатые полые образования, лишенные мембраны. Внешний диаметр составляет 24 нм, ширина просвета – 15 нм, толщина стенки – около 5 нм. В свободном состоянии представлены в цитоплазме, также являются структурными элементами жгутиков, цент-риолей, веретена деления, ресничек. Микротрубочки построены из стереотипных белковых субъединиц путем их полимеризации. В любой клетке процессы полимеризации идут параллельно процессам деполимеризации. Причем соотношение их определяется количеством микротрубочек. Микротрубочки имеют различную устойчивость к разрушающим их факторам, например, к колхицину (это химическое вещество, вызывающее деполимеризацию). Функции микротрубочек: 1) являются опорным аппаратом клетки; 2) определяют формы и размеры клетки; 3) являются факторами направленного перемещения внутриклеточных структур. Микрофиламенты Это тонкие и длинные образования, которые обнаруживаются по всей цитоплазме. Иногда образуют пучки. Виды микрофиламентов: 1) актиновые. Содержат сократительные белки (актин), обеспечивают клеточные формы движения (например, амёбоидные), играют роль клеточного каркаса, участвуют в организации перемещений органелл и участков цитоплазмы внутри клетки; 2) промежуточные (толщиной 10 нм). Их пучки обнаруживаются по периферии клетки под плазмалеммой и по окружности ядра. Выполняют опорную (каркасную) роль. В разных клетках (эпителиальных, мышечных, нервных, фибробластах) построены из разных белков. Микрофиламенты, как и микротрубочки, построены из субъединиц, поэтому их количество определяется соотношением процессов полимеризации и деполимеризации.
Внутренняя среда клетки Внутри клетки находится цитоплазма. Она состоит из жидкой части – гиалоплазмы (матрикса), органелл и цитоплазматических включений. Гиалоплазма Гиалоплазма – основное вещество цитоплазмы, заполняет все пространство между плазматической мембраной, оболочкой ядра и другими внутриклеточными структурами. Гиалоплазму можно рассматривать как сложную коллоидную систему, способную существовать в двух состояниях: золеобразном (жидком) и гелеобразном, которые взаимно переходят одно в другое. В процессе этих переходов осуществляется определенная работа, затрачивается энергия. Гиалоплазма лишена какой-либо определенной организации. Химический состав гиалоплазмы: вода (90 %), минеральные ионы, белки (ферменты гликолиза, обмена сахаров, азотистых оснований, белков и липидов). Некоторые белки цитоплазмы образуют субъединицы, дающие начало таким органеллам, как центриоли, микрофиламенты. Функции гиалоплазмы: 1) образование истинной внутренней среды клетки, которая объединяет все органеллы и обеспечивает их взаимодействие; 2) поддержание определенной структуры и формы клетки, создание опоры для внутреннего расположения органелл; 3) обеспечение внутриклеточного перемещения веществ и структур; 4) обеспечение адекватного обмена веществ как внутри самой клетки, так и с внешней средой. Включения Это относительно непостоянные компоненты цитоплазмы. Среди них выделяют: 1) запасные питательные вещества, которые используются самой клеткой в периоды недостаточного поступления питательных веществ извне (при клеточном голоде), – капли жира, гранулы крахмала или гликогена; 2) продукты, которые подлежат выделению из клетки, например, гранулы зрелого секрета в секреторных клетках (молоко в лактоцитах молочных желез); 3) балластные вещества некоторых клеток, которые не выполняют какой-либо конкретной функции (некоторые пигменты, например, липофусцин стареющих клеток).
Метаболизм Материальная сущность жизни проявляется, прежде всего, в непрерывном обмене веществ и энергии, который происходит между живой системой (клеткой, организмом, биоценозом) и окружающей его внешней средой. В этом смысле биологические системы являются открытыми. Разные организмы потребляют разные виды энергии, в связи с чем их делят на автотрофные и гетеротрофные. Автотрофные организмы (самопитающиеся) способны поглощать энергию неживой природы. Прежде всего, это зеленые растения, а также бурые и красные водоросли, использующие солнечный свет для процесса фотосинтеза – образования органического вещества глюкозы из неорганических воды и углекислого газа. К автотрофам относятся также сине-зеленые водоросли (цианеи) и некоторые бактерии, способные к реакциям хемосинтеза – синтеза органических веществ за счет энергии простых химических реакций. При этом первичная энергия (солнечная или химическая) преобразуется в энергию химических связей сложных органических молекул, так что автотрофы как бы сами создают себе пищу. Гетеротрофные организмы (питающиеся за счет других) – человек, все животные, грибы, а также многие бактерии, – получают пищу в виде готовых органических веществ, произведенных автотрофами, в основном растениями. В составе этой пищи они получают и энергию, заключенную в химических связях. Если органическое вещество пищи расщепить на более простые вещества, освобождается энергия. По существу гетеротрофы получают ту же солнечную энергию, но преобразованную зелеными растениями в химическую. Отсюда ясна огромная роль растительных организмов как посредника в энергетическом обеспечении животных и человека. Избавиться от этой зависимости, получать какую-либо энергию прямо из неживой природы человечество еще не научилось. И хотя академик В. И. Вернадский выдвигал такую научную задачу, дальше фантастических произведений дело не продвинулось и вряд ли продвинется в обозримом будущем. Поэтому для биологов всего мира одной из приоритетных задач остается понять во всех деталях механизм фотосинтеза, чтобы максимально интенсифицировать его в растениях и по возможности воспроизвести в искусственных условиях.
Структура АТФ и её изменение в ходе метаболизма Реакции энергетического обмена. Независимо от исходного источника энергии все организмы, как автотрофы, так и гетеротрофы, сначала переводят энергию в удобное для дальнейшего использования состояние. Это так называемые макроэргические (богатые энергией) связи в молекулах аденозинтрифосфорной кислоты – АТФ. Образуются молекулы АТФ из аденозиндифосфорной (АДФ) или аденозинмонофосфорной (АМФ) кислоты и свободных молекул фосфорной кислоты, но при непременном поглощении внешней энергии – солнечной или химической (эндотермическая реакция). Количество энергии, запасенное в макроэргической связи, на порядок больше, чем в обычных связях, например, внутри молекулы глюкозы, поэтому в составе АТФ энергию удобно хранить и транспортировать в пределах клетки. В местах потребления этой энергии АТФ распадается на АДФ и фосфат (при крайней необходимости даже на АМФ и два фосфата), а освобожденная энергия расходуется на ту или иную работу – синтез глюкозы в хлоропластах растительных клеток, синтез белков и других макромолекул, транспорт веществ в клетку и из клетки, движение и др. АДФ (АМФ) и фосфат могут снова соединиться, захватив очередную порцию внешней энергии, а потом разрушиться и отдать энергию в работу. Циклические преобразования АТФ многократно повторяются. Таким образом, АТФ выступает в качестве универсального переносчика энергии внутри клетки, своеобразной разменной монетой в энергетических платежах за внутриклеточные процессы.
Пути анаболизма и катаболизма в клетке Проблема клеточной энергетики сводится к пониманию первичных источников энергии и механизмов ее перевода в АТФ. В общем виде ситуация такова: у фотосинтетических аутотрофных организмов синтез АТФ из АДФ и фосфата генерируется солнечной энергией, у гетеротрофов – энергией от окисления пищевых продуктов. Таким образом, растениям для синтеза АТФ нужен свет, животным и человеку нужна органическая пища. Свет является первичным источником энергии, он используется в реакциях фотосинтезау растений. По конечной сути реакция фотосинтеза довольно проста: 6СО2 + 6H2O + энергия света → С6Н12О6 + 6О2↑ С помощью энергии света из углекислого газа и воды синтезируется 6-углеродное органическое вещество - глюкоза (моносахарид), и в качестве «лишнего» продукта образуется кислород, который уходит в атмосферу. На самом деле эта реакция более сложная, она состоит из двух стадий: световой и темновой. Сначала на свету с помощью особого Mg-содержащего пигмента хлорофилла вода расщепляется на кислород и водород, а энергия водорода передается на синтез АТФ. Только потом, в темновой стадии, водород соединяется с углекислым газом и образуется глюкоза. При этом часть АТФ расщепляется, отдавая энергию глюкозе. Глюкоза вместе с минеральными веществами, поступающими в растение из почвы (соли азота, серы, фосфора, железа, магния, кальция, калия, натрия и др.), становится основой для более сложных синтезов – образуются полисахариды, липиды, белки, нуклеиновые кислоты, из которых строятся рабочие структуры клеток. Но и эти синтезы, как и синтез глюкозы, требуют энергетических затрат. Прямое использование света здесь невозможно (эволюция не создала таких энергетических переходов), поэтому некоторая часть глюкозы тратится как энергетический субстрат, то есть глюкоза становится вторичным источником энергии. Глюкоза расщепляется и отдает энергию – сначала на синтез АТФ, а после расщепления АТФ – на биосинтезы макромолекул. Значительная часть АТФ, как уже сказано выше, расходуется на другую работу – транспорт веществ, движение клетки и др. Наиболее эффективно глюкоза расщепляется с участием кислорода: C6H12O6 + 6O2 → 6CO2 + 6H2O + энергия С химической точки зрения это полное окисление – «горение» глюкозы. В живой клетке «горение» происходит замедленно, поэтапно, так что энергия выделяется малыми порциями, и большая ее часть (около 55 %) используется на синтез АТФ, остальная рассеивается в виде тепла. Полное окисление одной молекулы глюкозы обеспечивает синтез 38 молекул АТФ. Поскольку кислород для окисления мы вдыхаем с атмосферным воздухом, то и на химическом уровне окисление глюкозы кислородом называют дыханием. Главная черта растительной автотрофной клетки – способность к фотосинтезу, который обеспечивает первый этап построения органического вещества, в форме глюкозы. Но и дыхание в полной мере присуще растениям, так как именно этот процесс извлекает энергию из глюкозы (а также из жиров и лишних белков), переводит ее временно в АТФ и далее в сложные макромолекулы. Эта же схема, но с изъятием реакции фотосинтеза, соответствует и гетеротрофному метаболизму животных клеток. В этом случае глюкоза (а также другие углеводы, жиры, трофические белки и др.) поступают в клетку извне в готовом виде. Часть этих материалов идет на дыхание (в топку, для извлечения энергии через синтез АТФ), а часть, после некоторой переделки, на синтез новых макромолекул как строительный материал. Таким образом, пища у гетеротрофов (то есть и у нас с вами) имеет двойное назначение – энергетическое и пластическое (строительное). Между пластическим обменом (анаболизмом) и энергетическим (катаболизмом) существует неразрывноеединство. Энергия поглощается из внешней среды, преобразуется в АТФ, прежде всего, для осуществления строительных процессов, для построения живой материи. А построение живой материи, то есть синтез макромолекул из простых неорганических веществ, возможен только с поглощением внешней энергии. Неклеточные формы жизни Вирусы – доклеточные формы жизни, которые являются облигатными внутриклеточными паразитами, т. е. могут существовать и размножаться только внутри организма хозяина. Вирусы были открыты Д. И. Ивановским в 1892 г. (он изучал вирус табачной мозаики), но доказать их существование удалось намного позднее. Многие вирусы являются возбудителями заболеваний, таких как СПИД, коревая краснуха, эпидемический паротит (свинка), ветряная и натуральная оспа. Вирусы имеют микроскопические размеры, многие из них способны проходить через любые фильтры. В отличие от бактерий, вирусы нельзя выращивать на питательных средах, так как вне организма они не проявляют свойств живого. Вне живого организма-хозяина вирусы представляют собой кристаллы веществ, не имеющих никаких свойств живых систем.
Схема строения бактериофага Зрелые вирусные частицы называются вирионами. Фактически они представляют собой геном, покрытый сверху белковой оболочкой. Эта оболочка – капсид. Она построена из белковых молекул, защищающих генетический материал вируса от воздействия нуклеаз – ферментов, разрушающих нуклеиновые кислоты. У некоторых вирусов поверх капсида располагается суперкапсидная оболочка, также построенная из белка. Генетический материал представлен нуклеиновой кислотой. У одних вирусов это ДНК (так называемые ДНК-вирусы), у других – РНК (РНК-вирусы). РНК-вирусы также называют ретровирусами, так как для синтеза вирусных белков в этом случае необходима обратная транскрипция, которая осуществляется ферментом – обратной транскриптазой (ревертазой) и представляет собой синтез ДНК на базе РНК.
Вирус иммунодефицита человека – ВИЧ
Бактериофаги – паразиты бактерий Размножение вирусов При внедрении вируса внутрь клетки-хозяина происходит освобождение молекулы нуклеиновой кислоты от белка, поэтому в клетку попадает только чистый и незащищенный генетический материал. Если вирус ДНК, то молекула ДНК встраивается в молекулу ДНК хозяина и воспроизводится вместе с ней. Так появляются новые вирусные ДНК, неотличимые от исходных. Все процессы, протекающие в клетке, замедляются, клетка начинает работать на воспроизводство вируса. Так как вирус является облигатным паразитом, то для его жизни необходима клетка-хозяин, поэтому она не погибает в процессе размножения вируса. Гибель клетки происходит только после выхода из нее вирусных частиц. Если это ретровирус, внутрь клетки-хозяина попадает его РНК. Она содержит гены, обеспечивающие обратную транскрипцию: на матрице РНК строится одноцепочечная молекула ДНК. Из свободных нуклеотидов достраивается комплементарная цепь, которая и встраивается в геном клетки-хозяина. С полученной ДНК информация переписывается на молекулу и-РНК, на матрице которой затем синтезируются белки ретровируса. Бактериофаги Это вирусы, паразитирующие на бактериях. Они играют большую роль в медицине и широко применяются при лечении гнойных заболеваний, вызванных стафилококками и др. Бактериофаги имеют сложное строение. Генетический материал находится в головке бактериофага, которая сверху покрыта белковой оболочкой. В центре головки находится атом магния. Далее идет полый стержень, который переходит в хвостовые нити. Их функция – узнавать свой вид бактерий, осуществлять прикрепление фага к клетке. После прикрепления ДНК выдавливается в бактериальную клетку, а оболочки остаются снаружи.
Хромосомы
Наследственность и изменчивость в живой природе существуют благодаря хромосомам, генам, дезоксирибонуклеиновой кислоте (ДНК). Исследование строения хромосом началось после их открытия и описания в XIX веке. Эти тельца и нити содержатся в прокариотических организмах (безъядерных) и эукариотических клетках (в ядрах). Изучение под микроскопом позволило установить, что такое хромосома с морфологической точки зрения. Это подвижное нитевидное тельце, которое различимо в определенные фазы клеточного цикла. В интерфазе весь объем ядра занимает хроматин. В другие периоды различимы хромосомы в виде одной или двух хроматид. Лучше видны эти образования во время клеточных делений — митоза или мейоза. В эукариотических клетках чаще можно наблюдать крупные хромосомы линейного строения. У прокариотов они меньше, хотя есть исключения. Клетки зачастую включают более одного типа хромосом, например свои собственные небольшие «частицы наследственности» есть в митохондриях и хлоропластах. Каждая хромосома обладает индивидуальным строением, отличается от других особенностями окрашивания. При изучении морфологии важно определить положение центромеры, длину и размещение плеч относительно перетяжки. В набор хромосом обычно входят следующие формы: метацентрические, или равноплечие, для которых характерно срединное расположение центромеры; субметацентрические, или неравноплечие (перетяжка смещена в сторону одного из теломеров); акроцентрические, или палочковидные, в них центромера находится практически на конце хромосомы; точковые с трудно поддающейся определению формой. Хромосомы состоят из генов — функциональных единиц наследственности. Теломеры — концы плеч хромосомы. Эти специализированные элементы служат для защиты от повреждения, препятствуют слипанию фрагментов. Центромера выполняет свои задачи при удвоении хромосом. На ней есть кинетохор, именно к нему крепятся структуры веретена деления. Каждая пара хромосом индивидуальна по месту расположения центромеры. Нити веретена деления работают таким образом, что в дочерние клетки отходит по одной хромосоме, а не обе. Равномерное удвоение в процессе деления обеспечивают точки начала репликации. Дупликация каждой хромосомы начинается одновременно в нескольких таких точках, что заметно ускоряет весь процесс деления. Общее число хромосом, их особенности — характерный признак вида. У мухи-дрозофилы их количество — 8, у шимпанзе — 48, у человека — 46. Это число является постоянным для клеток организмов, которые относятся к одному виду. Для всех эукариотов существует понятие «диплоидные хромосомы». Это полный набор, или 2n, в отличие от гаплоидного — половинного количества (n). Хромосомы в составе одной пары гомологичны, одинаковы по форме, строению, местоположению центромер и других элементов. Гомологи имеют свои характерные особенности, которые их отличают от других хромосом в наборе. Окрашивание основными красителями позволяет рассмотреть, изучить отличительные черты каждой пары. Диплоидный набор хромосом присутствует в соматических клетках, гаплоидный же — в половых (так называемых гаметах). У млекопитающих и других живых организмов с гетерогаметным мужским полом формируются два вида половых хромосом: Х-хромосома и Y. Самцы обладают набором XY, самки — XX. Изменение числа, формы или отдельных участков хромосом – мутации, приводит к искажению наследственной информации, и, как следствие, к заболеваниям, уродствам или летальному исходу. Клетки организма человека содержат 46 хромосом. Все они объединяются в 23 пары, составляющие набор. Есть два типа хромосом: аутосомы и половые. Первые образуют 22 пары — общие для женщин и мужчин. От них отличается 23-я пара — половые хромосомы, которые в клетках мужского организма являются негомологичными. Генетические черты связаны с половой принадлежностью. Для их передачи служат Y и Х-хромосома у мужчин, две X у женщин. Аутосомы содержат оставшуюся часть информации о наследственных признаках. Существуют методики, позволяющие индивидуализировать все 23 пары. Они хорошо различимы на рисунках, когда окрашены в определенный цвет.
Кариотип человека Половое размножение Эволюционный смысл полового размножения Половое размножение встречается в основном у высших организмов. Это более поздний вид размножения (существует около 3 млрд лет). Оно обеспечивает значительное генетическое разнообразие и, следовательно, большую фенотипическую изменчивость потомства; организмы получают большие эволюционные возможности, возникает материал для естественного отбора. Помимо полового размножения, существует половой процесс. Суть его в том, что обмен генетической информацией между особями происходит, но без увеличения числа особей. Формированию гамет у многоклеточных предшествует мейоз. Половой процесс состоит в объединении наследственного материала от двух разных источников (родителей). При половом размножении потомство генетически отличается от своих родителей, так как между родителями происходит обмен генетической информацией. Основой полового размножения является мейоз. Родителями являются две особи – мужская и женская, они вырабатывают разные половые клетки. В этом проявляется половой диморфизм, который отражает различие задач, выполняемых при половом размножении мужским и женским организмами. Половое размножение осуществляется через гаметы – половые клетки, имеющие гаплоидный набор хромосом и вырабатывающиеся в родительских организмах. Слияние родительских клеток приводит к образованию зиготы, из которой в дальнейшем образуется организм-потомок. Половые клетки образуются в гонадах – половых железах (в яичниках у самок и семенниках у самцов). Процесс образования половых клеток называется гаметогенезом (овогенезом у самок и сперматогенезом у самцов). Если мужские и женские гаметы образуются в организме одной особи, то ее называют гермафродитной. Гермафродитизм бывает истинный (особь имеет гонады обоих полов) и ложный гермафродитизм (особь имеет половые железы одного типа – мужского или женского, а наружные половые органы и вторичные половые признаки обоих полов). Виды полового размножения У одноклеточных организмов выделяют две формы полового размножения – копуляцию и конъюгацию. При конъюгации (например, у инфузорий) специальные половые клетки (половые особи) не образуются. У этих организмов имеются два ядра – макро- и микронуклеус. Обычно инфузории размножаются делением надвое. При этом микронуклеус сначала делится митотически. Из него формируются стационарное и мигрирующее ядра, имеющие гаплоидный набор хромосом. Затем две клетки сближаются, между ними образуется протоплазматический мостик. По нему происходит перемещение в цитоплазму партнера мигрирующего ядра, которое затем сливается со стационарным. Формируются обычные микро- и макронуклеусы, клетки расходятся. Так как при этом процессе не происходит увеличения количества особей, то говорят о половом процессе, а не о половом размножении. Однако происходит обмен (рекомбинация) наследственной информацией, поэтому потомки генетически отличаются от своих родителей. При копуляции (у простейших) происходят образование половых элементов и их попарное слияние. При этом две особи приобретают половые различия и полностью сливаются, образуя зиготу. Происходят объединение и рекомбинация наследственного материала, поэтому особи генетически отличны от родительских. Различия между гаметами В процессе эволюции степень различия гамет нарастает. Сначала имеет место простая изогамия, когда половые клетки еще не имеют дифференцировки. При дальнейшем усложнении процесса возникает анизогамия: мужские и женские гаметы различаются, однако не качественно, а количественно (у хламидомонад). Наконец, у водоросли вольвокса большая гамета становится неподвижной и самой крупной из всех гамет. Такая форма анизогамии, когда гаметы резко различны, называется оогамией. У многоклеточных животных (в том числе у человека) имеет место исключительно оогамия. Среди растений изогамия и анизогамия встречаются только у водорослей. |
||
|
|
Последнее изменение этой страницы: 2018-05-10; просмотров: 727. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |