|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
НЕЙРОН. ЕГО СТРОЕНИЕ И ФУНКЦИИСтр 1 из 108Следующая ⇒ Юрий Александров Основы психофизиологии - Александров Ю.И. (ред.)
«Александров Ю.И. (ред.) - Основы психофизиологии»: Инфра-М; Москва; 1998 Аннотация
В учебнике «Основы психофизиологии» раскрыты все темы, составляющие в соответствии с Государственным образовательным стандартом высшего профессионального образования содержание курса по психофизиологии, и дополнительно те вопросы, которые представляют собой «точки роста» и привлекают значительное внимание исследователей. В учебнике описаны основные методологические подходы и методы, разработанные как в отечественной, так и в зарубежной психофизиологии, последние достижения этой науки. Настоящий учебник, который отражает современное состояние психофизиологии во всей её полноте, предназначен студентам, аспирантам, научным сотрудникам, а также всем тем, кто интересуется методологией науки, психологией, психофизиологией, нейронауками, методами и результатами объективного изучения психики.
Основы психофизиологии: Учебник / Отв. ред. Ю.И. Александров. - М.: ИНФРА-М, 1997
ПРЕДИСЛОВИЕ
По-видимому, все, кто откроет эту книгу, согласятся со следующим высказыванием И.П. Павлова: «В сущности, интересует нас в жизни только одно: наше психическое содержание» [1949, с. 351]. В то же время поддержка выдвинутого ещё Аристотелем положения: «Исследование души есть дело естествоиспытателя» [Аристотель, 1937, с. 7], не будет столь же безоговорочной. Самое общее уточнение могло бы состоять в том, что познание «психического содержания» – дело не только науки, но и других видов человеческой деятельности, таких, например, как искусство или религия. Если же рассмотреть лишь один вид деятельности – науку, то и здесь оказывается, что «психическое содержание» исследуется представителями как естественных, например физиологии, так и общественных наук, к которым принято относить психологию, сочетающую естественнонаучные методы с «герменевтическими» (моделирование в психике исследователя психики испытуемого, зависящее от индивидуально-психологических особенностей исследователя [Дружинин, 1993]).  Контакты между названными науками, которые возникают при решении проблем, представляющих взаимный интерес, часто «искрят» [Швырков, 1995], что вызывает у многих физиологов и психологов желание изолировать свою дисциплину, оградить её от посторонних посягательств. Однако выдающимся психологам уже давно было очевидно, что предпринимаемые как психологами, так иногда и физиологами попытки эмансипировать психологию от физиологии совершенно неправомерны, поскольку предмет психологии – нейропсихический процесс [Бехтерев, 1991], целостная психофизиологическая реальность [Выготский, 1982], которая лежит в основе всех без исключения психических процессов, включая и самые высшие [Рубинштейн, 1973]. Со стороны психофизиологии также были приведены веские аргументы в пользу того, что самостоятельная, отделённая от психологии физиология не может выдвинуть обоснованной концепции целостной деятельности мозга [Швырков, 1995]. «Изоляция какой-либо дисциплины есть верный показатель её ненаучности», – справедливо заключает М. Бунге, отвечая на вопрос: «Является ли психология автономной дисциплиной?» Психология же тесно взаимодействует и даже перекрывается с биологией, в частности, с физиологией [Bunge, 1990], причём область их взаимодействия постоянно увеличивается. Логика развития методологии и методов науки, а также «социальные заказы», заставляющие преодолевать междисциплинарные барьеры [Абульханова и др., 1996], определяют возможность и необходимость всё большего привлечения методов физиологии для разработки проблем профессионального и психического здоровья, сознания и бессознательного, изучения структуры сложной деятельности человека – совместной, речевой, операторской и мн. др. Связь и взаимозависимость психологии и физиологии настолько сильны, что позволяют рассматривать их развитие как коэволюцию. Вкратце охарактеризуем этот процесс. Современная психология в значительной степени представлена интуитивной бытовой, или «обыденной психологией» (folk psychology), под которой понимается основанное на здравом смысле, не требующее точных определений понимание психических процессов и состояний [Churchland, 1986]. Дело в том, что такие понятия обыденной психологии, как память, внимание, воля, ум, влечение, чувство и другие не только употребляются в быту для объяснения и предсказания поведения людей, но и влияют на формирование собственно научного знания [Sternberg, 1985; Semin, 1987]. Они используются в психологических исследованиях, как при обосновании проблем исследования, так и при трактовке его результатов. Закономерности и феномены, выявляемые в подобных исследованиях, становятся базой не только для следующих психологических исследований, но и для формулировки задач экспериментов, в которых применяются методы физиологии. Решение этих задач способствует пересмотру и фрагментации исходных концепций и понятий (сколько, например, разнообразнейших процессов, «систем» и прочего объединяется сейчас термином «память»!), формулировке новых вопросов и т.д. В конце концов в ряде случаев может даже оказаться, что выяснять надо что-то совсем другое. Скажем, современная физиология не исследует, как образуются и движутся «животные духи». Наряду с обыденной психологией существует и обыденная физиология, которая взаимодействует и с обыденной психологией, и с собственно наукой. Однако здесь мы можем пренебречь самостоятельным значением обыденной физиологии и ограничиться рассмотрением лишь обыденной психологии, полагая, что последняя инкорпорирует ту часть обыденного физиологического знания, которая имеет отношение к интересующему нас проблемному полю. Следует подчеркнуть, что в процессе коэволюции не происходит «истребления» психологии [Ярошевский, 1996], её исчезновения, замены физиологией, так как психологическое исследование формирует специфический компонент описания поведения и деятельности, необходимый как для самой психологии, так и для сопредельных дисциплин. Происходит же постепенное замещение бытовых понятий в психологии и физиологии научными. В связи с этим предполагается, что по ходу коэволюции психология и физиология будут всё меньше зависеть от обыденной психологии и на определённом этапе замещение завершится [Churchland, 1986]. По-видимому, это предположение полностью справедливо лишь в том случае, если говорить не об обыденной психологии вообще, а о ныне существующей обыденной психологии. В процессе развития на место замещённых придут новые понятия обыденной науки. На чём основано это утверждение? Ещё до возникновения науки, в доисторическую эпоху, представления о психике, или душе, складывались у людей как обобщённые характеристики внешнего поведения, как гипотезы о его детерминантах и механизмах. В связи с этим даже в наиболее примитивных языках имеются обозначения психических свойств и состояний (см. в [Швырков, 1995]). С появлением науки она становится, наряду с религией, искусством, обыденным опытом и т.д., одним из источников концепций, идей и терминов, включающихся в обыденное сознание и формирующих обыденную науку. Люди начинают рассматривать их как само собой разумеющиеся и составляющие «реальность» [Московичи, 1995]. Можно полагать в связи с этим, что обыденная наука продолжит своё существование на всём протяжении процесса коэволюции, осуществляя «обмен» с собственно наукой. Первая будет поставщиком концепций и проблем (конечно, не исключительным – много проблем в собственно науке имеет внутринаучное происхождение) мировоззренческого или практического характера, вторая будет решать эти проблемы и возвращать переработанные концепции, внедрять новые или устранять дискредитированные. При этом количество ассимилированных и преобразованных понятий собственно науки в науке обыденной будет постоянно и быстро увеличиваться, если авторитет науки сохранится и общество не последует антисциентистским рекомендациям, таким, как дополнить отделение государства от церкви отделением его и от науки [Фейрабенд, 1986]. Каково же место психофизиологии, науки, обязанной своим происхождением и даже названием сосуществованию психологии и физиологии и призванной установить между ними связь, в описанной ранее коэволюции? Каков её специфический вклад? Можно ли свести роль психофизиологии к использованию методов физиологии для изучения психических процессов и состояний? Ответы, которые дают на эти вопросы разные авторы, в том числе и авторы настоящего учебника, значительно различаются. Известно, что даже физическую систему нельзя описать каким-либо одним теоретическим языком, множественность точек зрения на неё неустранима [Пригожий, Стенгерс, 1986]. Тем более не должно вызывать удивления существование различающихся позиций, разных теоретических языков и школ в психофизиологии, которая, по-видимому, имеет дело с наиболее сложным комплексом проблем, стоящих перед человеком. Упомянутые различия находят своё выражение не только в многообразии ответов на один и тот же вопрос, но и в том, что отдельные вопросы психофизиологии, рассматриваемые одними исследователями как центральные, с других теоретических позиций могут расцениваться как малозначимые или даже неверно поставленные. Поэтому мы полагали, что в учебнике по психофизиологии, написанном одним автором, даже сам набор освещаемых проблем окажется в сильнейшей мере зависимым от взглядов автора. Кроме того, мы учитывали, что психофизиология находится на стыке разных наук: философии, психологии, нейронаук, физиологии, в том числе физиологии высшей нервной деятельности, генетики, биохимии и т.д. Во многом ситуация здесь сходна с имевшей место при подготовке руководства по экспериментальной психологии [I960], в предисловии к которому С.С.Стивенс отмечал, что никто из представителей этой науки не обладает достаточной эрудицией, чтобы справиться с подобной задачей в одиночку. В связи с уже сказанным мы, считая, что высокое качество всех разделов учебника, а также достаточно полный охват обширного проблемного поля психофизиологии может обеспечить только совместная деятельность коллектива учёных, обладающих взаимно дополняющей квалификацией и представляющих позиции разных научных школ, выбрали коллективный путь его подготовки. Список глав настоящего учебника включает все пункты Государственного образовательного стандарта высшего профессионального образования (М., 1995), а также дополнительно и те вопросы, которые представляют собой «точки роста», привлекают значительное внимание исследователей и игнорирование которых не позволило бы считать учебник отражающим современное состояние психофизиологии. В результате содержание учебника соответствует не только Государственному образовательному стандарту высшего профессионального образования, но и Программе по психофизиологии, подготовленной Советом по психологии Объединения государственных университетов Российской Федерации (М., 1996). Глава 1 подготовлена канд. мед. наук Б.Н. Безденежных (параграф 1) и докт. психол. наук Т.Н. Греченко (параграф 2), главы 2 и 9 – канд. мед. наук Б.Н. Безденежных, главы 3 и 4 – докт. биол. наук, проф. И.А. Шевелевым, глава 5 – докт. мед. наук, акад. B. C. Гурфинкелем и канд. биол. наук Ю.С. Левиком, глава 6 – докт. психол. наук Т.Н. Греченко (параграфы 1–5) и докт. биол. наук, проф. А.Н. Лебедевым (параграфы 6 – 8), глава 7 – докт. мед. наук, акад. П.В. Симоновым, главы 8 и 10 – докт. психол. наук, проф. Н.Н. Даниловой, глава 11 – докт. мед. наук, проф. A.M. Иваницким, глава 12 – докт. мед. наук, проф. Э.А. Костандовым, глава 13 – канд. мед. наук Д.Г. Шевченко (параграф 8 – совместно с докт. психол. наук Ю.И. Александровым), глава 14 – докт. психол. наук Ю.И. Александровым, глава 15 – канд. психол. наук А.Г. Горкиным, глава 16 – канд. психол. наук И.О. Александровым и канд. психол. наук Н.Е. Максимовой, глава 17 – канд. психол. наук М.В. Бодуновым, глава 18 – канд. психол. наук Л.Г. Дикой, глава 19 – канд. психол. наук В.В. Гавриловым. Настоящий учебник, который отражает современное состояние психофизиологии во всей её полноте, предназначен студентам, аспирантам, научным сотрудникам, а также всем тем, кто интересуется методологией науки, психологией, психофизиологией, нейронауками, методами и результатами объективного изучения психики. Доктор психол. наук Ю.И. Александров
Глава 1 МОЗГ
ОБЩИЕ СВЕДЕНИЯ
Традиционно со времён французского физиолога Биша (начало XIX в.) нервную систему разделяют на соматическую и вегетативную, в каждую из которых входят структуры головного и спинного мозга, называемые центральной нервной системой (ЦНС), а также лежащие вне спинного и головного мозга и поэтому относящиеся к периферической нервной системе нервные клетки и нервные волокна, иннервирующие органы и ткани организма. Соматическая нервная система представлена эфферентными (двигательными) нервными волокнами, иннервирующими скелетную мускулатуру, и афферентными (чувствительными) нервными волокнами, идущими в ЦНС от рецепторов. Вегетативная нервная система включает в себя эфферентные нервные волокна, идущие к внутренним органам и рецепторам, и афферентные волокна от рецепторов внутренних органов. По морфологическим и функциональным особенностям вегетативная нервная система разделяется на симпатическую и парасимпатическую. По своему развитию, а также структурной и функциональной организации нервная система человека имеет сходство с нервной системой разных видов животных, что существенно расширяет возможности её исследования не только морфологами и нейрофизиологами, но и психофизиологами. У всех видов позвоночных нервная система развивается из пласта клеток на наружной поверхности эмбриона – эктодермы. Часть эктодермы, называемая нервной пластинкой, сворачивается в полую трубку, из которой формируются головной и спинной мозг. В основе этого формирования лежит интенсивное деление эктодермальных клеток и формирование нервных клеток. Каждую минуту формируется примерно 250 000 клеток [Коуэн, 1982]. Молодые несформированные нервные клетки постепенно мигрируют из областей, где они возникли, к местам своей постоянной локализации и объединяются в группы. В результате стенка трубки утолщается, сама трубка начинает трансформироваться, и на ней появляются идентифицируемые участки мозга, а именно: в её передней части, которая будет в дальнейшем заключена в череп, образуются три первичных мозговых пузыря – это rhombencephalon, или задний мозг; mesencephalon, или средний мозг, и prosencephalon, или передний мозг (рис. 1.1 А, Б). Из задней части трубки формируется спинной мозг. Мигрировав на место постоянной локализации, нейроны начинают дифференцироваться, у них появляются отростки (аксоны и дендриты) и их тела приобретают определённую форму (см. параграф 2). Одновременно происходит дальнейшая дифференциация мозга. Задний мозг дифференцируется на продолговатый мозг, мост и мозжечок; в среднем мозге нервные клетки группируются в виде двух пар крупных ядер, называемых верхними и нижними бугорками четверохолмия. Центральное скопление нервных клеток (серое вещество) на этом уровне носит название покрышек среднего мозга. В переднем мозге происходят наиболее существенные изменения. Из него дифференцируются правая и левая камеры. Из выпячиваний этих камер в дальнейшем формируются сетчатки глаз. Остальная, большая часть, правой и левой камер превращается в полушария; эта часть мозга называется конечным мозгом (telencephalon), и наиболее интенсивное развитие она получает у человека. Образовавшийся после дифференциации полушарий центральный отдел переднего мозга получил название промежуточного мозга (diencephalon); он включает в себя таламус и гипоталамус с железистым придатком, или гипофизарным комплексом. Части мозга, расположенные ниже конечного мозга, т.е. от промежуточного до продолговатого мозга включительно, называют стволом мозга. Под влиянием сопротивления черепа интенсивно увеличивающиеся стенки конечного мозга отодвигаются назад и прижимаются к стволу мозга (рис. 1.1 В). Наружный слой стенок конечного мозга становится корой больших полушарий, а их складки между корой и верхней частью ствола, т.е. таламусом, образуют базальные ядра – полосатое тело и бледный шар. Кора больших полушарий мозга – это наиболее позднее в эволюции образование. По некоторым данным у человека и у других приматов не менее 70% всех нервных клеток ЦНС локализовано в коре больших полушарий [Наута, Фейртаг, 1982]; её площадь увеличена за счёт многочисленных извилин. В нижней части полушарий кора подворачивается вовнутрь и образует сложные складки, которые на поперечном срезе напоминают морского конька – гиппокамп.
Рис.1.1. Развитие мозга млекопитающих [Милнер, 1973] А. Расширение переднего конца нервной трубки и образование трёх отделов головного мозга Б Дальнейшее расширение и разрастание переднего мозга В . Разделение переднего мозга на промежуточный мозг (таломус и гипоталамус), базальные ядра и кору больших полушарий. Показано относительное расположение этих структур: 1 – передний мозг (prosencephalon); 2 – средний мозг (mesencepholon); 3 – задний мозг (rhombencephalon); 4 – спинной мозг (medulla spinalis); 5– боковой желудочек (ventriculus lateralis); 6 – третий желудочек (ventriculus tertius); 7 – сильвиев водопровод (aqueductus cerebri); 8 – четвёртый желудочек (ventriculus quartus); 9 – полушария мозга (hemispherium cerebri); 10 – таламус (thalamus) и гиполамус (hypothalamus); 11– базальные ядра (nuclei basalis); 12 – мост (pons) (вентрально) и мозжечок (cerebellum)(дорсально); 13 – продолговатый мозг (medulla oblongata). В толще стенок дифференцирующихся структур мозга в результате агрегации нервных клеток формируются глубинные мозговые образования в виде ядер, формаций и субстанций, причём в большинстве областей мозга клетки не только агрегируют друг с другом, но и приобретают некоторую предпочтительную ориентацию. Например, в коре головного мозга большинство крупных пирамидных нейронов выстраиваются в ряд таким образом, что их верхние полюса с дендритами направлены к поверхности коры, а нижние полюса с аксонами – в направлении белого вещества. С помощью отростков нейроны формируют связи с другими нейронами; при этом аксоны многих нейронов, прорастая в отдалённые участки, образуют специфические анатомически и гистологически выявляемые проводящие пути. Следует отметить, что процесс формирования структур мозга и проводящих путей между ними происходит не только за счёт дифференциации нервных клеток и прорастания их отростков, но и за счёт обратного процесса, заключающегося в гибели некоторых клеток и ликвидации ранее сформированных связей. В результате описанных ранее трансформаций образуется мозг – предельно сложное морфологическое образование. Схематическое изображение мозга человека представлено на рис. 1.2.
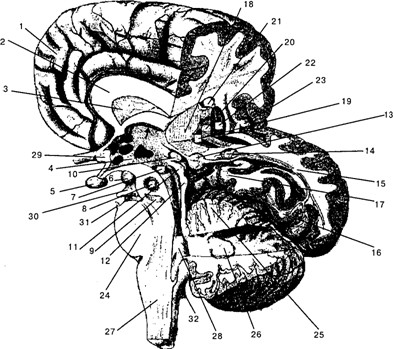
Рис. 1.2. Головной мозг (правое полушарие; частично удалены теменная, височная и затылочная области): 1 – медиальная поверхность лобной области правого полушария; 2 – мозолистое тело (corpus callosum); 3 – прозрачная перегородка (septum pellucidum); 4 – ядра гипоталамуса (nuclei hypothalami); 5 – гипофиз (hypophisis); 6 – мамилярное тело ( corpus mamillare); 7– субталамическое ядро (nucleus subthalamicus); 8 – красное ядро (nucleus ruber) (проекция); 9 – чёрная субстанция (substantia nigra)(проекция); 10– шишковидная железа (corpus pineale); 11 – верхние бугорки четверохолмия (colliculi superior tecti mesencepholi); 12 – нижние бугорки четверохолмия (colliculi inferior tecti mesencephali); 13 – медиальное коленчатое тело (МКТ) (corpus geniculatum mediale); 14 – латеральное коленчатое тело (ЛКТ) (corpus geniculatum laterale); 15 – нервные волокна, идущие от ЛКТ в первичную зрительную кору; 16 – шпорная извилина (sulcus calcarinus); 17– гиппокампальная извилина (girus hippocampalis); 18 – таламус (thalamus); 19 – внутренняя часть бледного шара (globus pallidus); 20 – наружная часть бледного шара; 21 – хвостатое ядро (nucleus caudatus); 22 – скорлупа (putamen); 23 – островок (insula); 24 – мост (pons); 25 – мозжечок (кора)(cerebellum); 26– зубчатое ядро мозжечка (nucleus dentatus); 27– продолговатый мозг (medulla oblongata); 28– четвёртый желудочек (ventriculus quartus); 29 – зрительный нерв (nervus opticus); 30 – глазодвигательный нерв (nervus oculomotoris); 31 – тригеминальный нерв (nervus trigeminus); 32 – вестибулярный нерв (nervus vestibularis). Стрелкой обозначен свод
НЕЙРОН. ЕГО СТРОЕНИЕ И ФУНКЦИИ
Мозг человека состоит из 10 12 нервных клеток. Обычная нервная клетка получает информацию от сотен и тысяч других клеток и передаёт сотням и тысячам, а количество соединений в головном мозге превышает 10 14 - 10 15 . Открытые более 150 лет тому назад в морфологических исследованиях Р. Дютроше, К. Эренберга и И. Пуркинье, нервные клетки не перестают привлекать к себе внимание исследователей. Как независимые элементы нервной системы они были открыты сравнительно недавно – в XIX в. Гольджи и Рамон-и-Кахал применили достаточно совершенные методы окраски нервной ткани и нашли, что в структурах мозга можно выделить клетки двух типов: нейроны и глию. Нейробиолог и нейроанатом Рамон-и-Кахал использовал метод окраски по Гольджи для картирования участков головного и спинного мозга. В результате была показана не только чрезвычайная сложность, но и высокая степень упорядоченности нервной системы. С тех пор появились новые методы исследования нервной ткани, позволяющие выполнить тонкий анализ её строения, – например, использование гисторадиохимии выявляет сложнейшие связи между нервными клетками, что позволяет выдвигать принципиально новые предположения о построении нейронных систем. Имеющая исключительно сложное строение, нервная клетка – это субстрат самых высокоорганизованных физиологических реакций, лежащих в основе способности живых организмов к дифференцированному реагированию на изменения внешней среды. К функциям нервной клетки относят передачу информации об этих изменениях внутри организма и её запоминание на длительные сроки, создание образа внешнего мира и организацию поведения наиболее целесообразным способом, обеспечивающим живому существу максимальный успех в борьбе за своё существование. Исследования основных и вспомогательных функций нервной клетки в настоящее время развились в большие самостоятельные области нейробиологии. Природа рецепторных свойств чувствительных нервных окончаний, механизмы межнейронной синаптической передачи нервных влияний, механизмы появления и распространения нервного импульса по нервной клетке и её отросткам, природа сопряжения возбудительного и сократительного или секреторного процессов, механизмы сохранения следов в нервных клетках – всё это кардинальные проблемы, в решении которых за последние десятилетия достигнуты большие успехи благодаря широкому внедрению новейших методов структурного, электрофизиологического и биохимического анализов.
Размер и форма
Размеры нейронов могут быть от 1 (размер фоторецептора) до 1000 мкм (размер гигантского нейрона у морского моллюска Aplysia) (см. [Сахаров, 1992]). Форма нейронов также исключительно разнообразна. Наиболее ясно форма нейронов видна при приготовлении препарата полностью изолированных нервных клеток. Нейроны чаще всего имеют неправильную форму. Существуют нейроны, напоминающие «листик» или «цветок». Иногда поверхность клеток напоминает мозг – она имеет «борозды» и «извилины». Исчерченность мембраны нейронов увеличивает её поверхность более чем в 7 раз. В нервных клетках различимы тело и отростки. В зависимости от функционального назначения отростков и их количества различают клетки монополярные и мультиполярные. Монополярные клетки имеют только один отросток – это аксон. Согласно классическим представлениям, у нейронов один аксон, по которому возбуждение распространяется от клетки. Согласно же наиболее новым результатам, полученным в электрофизиологических исследованиях с использованием красителей, которые могут распространяться от тела клетки и прокрашивать отростки, нейроны имеют более чем один аксон. Мультиполярные (биполярные) клетки имеют не только аксоны, но и дендриты. По дендритам сигналы от других клеток поступают в нейрон. Дендриты в зависимости от их локализации могут быть базальными и апикальными. Дендритное дерево некоторых нейронов чрезвычайно разветвлено, а на дендритах находятся синапсы – структурно и функционально оформленные места контактов одной клетки с другой. Какие клетки более совершенны – униполярные или биполярные? Униполярные нейроны могут быть определённым этапом в развитии биполярных клеток. В то же время у моллюсков, которые на эволюционной лестнице занимают далеко не верхний этаж, нейроны униполярные. Новыми гистологическими исследованиями показано, что даже у человека при развитии нервной системы клетки некоторых структур мозга из униполярных «превращаются» в биполярные. Подробное исследование онтогенеза и филогенеза нервных клеток убедительно показало, что униполярное строение клетки является вторичным явлением и что во время эмбрионального развития можно шаг за шагом проследить постепенное превращение биполярных форм нервных клеток в униполярные. Рассматривать биполярный или униполярный тип строения нервной клетки как признак сложности строения нервной системы вряд ли верно. Отростки-проводники придают нервным клеткам способность объединяться в нервные сети различной сложности, что является основой для создания из элементарных нервных клеток всех систем мозга. Для приведения в действие этого основного механизма и его использования нервные клетки должны обладать вспомогательными механизмами. Назначением одного из них является превращение энергии различных внешних воздействий в тот вид энергии, который может включить процесс электрического возбуждения. У рецепторных нервных клеток таким вспомогательным механизмом являются особые сенсорные структуры мембраны, позволяющие изменять её ионную проводимость при действии тех или иных внешних факторов (механических, химических, световых). У большинства других нервных клеток – это хемочувствительные структуры тех участков поверхностной мембраны, к которым прилежат окончания отростков других нервных клеток (постсинаптические участки) и которые могут изменять ионную проводимость мембраны при взаимодействии с химическими веществами, выделяемыми нервными окончаниями. Возникающий при таком изменении локальный электрический ток является непосредственным раздражителем, включающим основной механизм электрической возбудимости. Назначение второго вспомогательного механизма – преобразование нервного импульса в процесс, который позволяет использовать принесённую этим сигналом информацию для запуска определённых форм клеточной активности.
Цвет нейронов
Следующая внешняя характеристика нервных клеток – это их цвет. Он также разнообразен и может указывать на функцию клетки – например, нейроэндокринные клетки имеют белый цвет. Жёлтый, оранжевый, а иногда и коричневый цвет нейронов объясняется пигментами, которые содержатся в этих клетках. Размещение пигментов в клетке неравномерно, поэтому её окраска различна по поверхности – наиболее окрашенные участки часто сосредоточены вблизи аксонного холмика. По-видимому, существует определённая взаимосвязь между функцией клетки, её цветом и её формой. Наиболее интересные данные об этом получены в исследованиях на нервных клетках моллюсков.
Синапсы
Биофизический и клеточно-биологический подход к анализу нейронных функций, возможность идентификации и клонирования генов, существенных для сигнализации, вскрыли тесную связь между принципами, которые лежат в основе синаптической передачи и взаимодействия клеток. В результате было обеспечено концептуальное единство нейробиологии с клеточной биологией. Когда выяснилось, что ткани мозга состоят из отдельных клеток, соединённых между собой отростками, возник вопрос: каким образом совместная работа этих клеток обеспечивает функционирование мозга в целом? На протяжении десятилетий споры вызывал вопрос о способе передачи возбуждения между нейронами, т.е. каким путём она осуществляется: электрическим или химическим. К середине 20-х гг. большинство учёных приняли ту точку зрения, что возбуждение мышц, регуляция сердечного ритма и других периферийных органов – результат воздействия химических сигналов, возникающих в нервах. Эксперименты английского фармаколога Г. Дейла и австрийского биолога О. Леви были признаны решающими подтверждениями гипотезы о химической передаче. Усложнение нервной системы развивается по пути установления связей между клетками и усложнения самих соединений. Каждый нейрон имеет множество связей с клетками-мишенями. Эти мишени могут быть нейронами разных типов, нейросекреторными клетками или мышечными клетками. Взаимодействие нервных клеток в значительной мере ограничено специфическими местами, в которые могут приходить соединения – это синапсы. Данный термин произошёл от греческого слова «застёгивать» и был введён Ч. Шеррингтоном в 1897 г. А на полвека раньше К. Бернар уже отмечал, что контакты, которые формируют нейроны с клетками-мишенями, специализированы, и, как следствие, природа сигналов, распространяющихся между нейронами и клетками-мишенями, каким-то образом изменяется в месте этого контакта. Критичные морфологические данные о существовании синапсов появились позже. Их получил С. Рамон-и-Кахал (1911), который показал, что все синапсы состоят из двух элементов – пресинаптической и постсинаптической мембраны. Рамон-и-Кахал предсказал также существование третьего элемента синапса – синаптической щели (пространства между пресинаптическим и постсинаптическим элементами синапса). Совместная работа этих трёх элементов и лежит в основе коммуникации между нейронами и процессами передачи синаптической информации. Сложные формы синаптических связей, формирующихся по мере развития мозга, составляют основу всех функций нервных клеток – от сенсорной перцепции до обучения и памяти. Дефекты синаптической передачи лежат в основе многих заболеваний нервной системы. Синаптическая передача через большую часть синапсов мозга опосредуется при взаимодействии химических сигналов, поступающих из пресинаптической терминали, с постсинаптическими рецепторами. В течение более чем 100 лет изучения синапса все данные рассматривались с точки зрения концепции динамической поляризации, выдвинутой С. Рамон-и-Кахалом. В соответствии с общепринятой точкой зрения синапс передаёт информацию только в одном направлении: информация течёт от пресинаптической к постсинаптической клетке, антероградно направленная передача информации обеспечивает финальный шаг в сформированных нейронных коммуникациях. Анализ новых результатов заставляет предполагать, что существенная часть информации передаётся и ретроградно – от постсинаптического нейрона к пресинаптическим терминалям нерва [Jessell, Kandel, 1993]. В некоторых случаях были идентифицированы молекулы, которые опосредуют ретроградную передачу информации. Это целый ряд веществ от подвижных маленьких молекул окиси азота до больших полипептидов, таких, как фактор роста нерва. Даже если сигналы, которые передают информацию ретроградно, различны по своей молекулярной природе, принципы, на основе которых эти молекулы действуют, могут быть сходными. Бидирекциональность передачи обеспечивается и в электрическом синапсе, в котором щель в соединительном канале формирует физическую связь между двумя нейронами, без использования нейромедиатора для передачи сигналов от одного нейрона на другой. Это позволяет осуществлять бидирекциональную передачу ионов и других маленьких молекул. Но реципрокная передача существует также в дендродендритных химических синапсах, где оба элемента имеют приспособления для высвобождения передатчика и ответа. Так как эти формы передачи часто трудно дифференцировать в сложных сетях мозга, случаев бидирекциональной синаптической коммуникации может оказаться значительно больше, чем это кажется сейчас. Бидирекциональная передача сигналов в синапсе играет важную роль в любом из трёх основных аспектов работы нервной сети: синаптической передаче, пластичности синапсов и созревании синапсов во время развития. Пластичность синапсов – это основа для формирования связей, которые создаются при развитии мозга и при научении. В обоих случаях требуется ретроградная передача сигналов от постк пресинаптической клетке, сетевой эффект которой заключается в том, чтобы сохранить или потенциировать активные синапсы. Ансамбль синапсов вовлекает координированное действие протеинов, высвобождаемых из преи постсинаптической клетки. Первичная функция белков состоит в том, чтобы индуцировать биохимические компоненты, требуемые для высвобождения передатчика из пресинаптической терминали, а также для того, чтобы организовать устройство для передачи внешнего сигнала постсинаптической клетке.
Электрическая возбудимость
Все функции, свойственные нервной системе, связаны с наличием у нервных клеток структурных и функциональных особенностей, обеспечивающих возможность генерации под влиянием внешнего воздействия особого сигнального процесса – нервного импульса (основными свойствами которого являются незатухающее распространение вдоль клетки, возможность передачи сигнала в необходимом направлении и воздействие с его помощью на другие клетки). Способность к генерации нервной клеткой распространяющегося нервного импульса определяется особым молекулярным устройством поверхностной мембраны, позволяющим воспринимать изменения проходящего через неё электрического поля, изменять практически мгновенно свою ионную проводимость и создавать за счёт этого трансмембранный ионный ток, используя в качестве движущей силы постоянно существующие между внеи внутриклеточной средой ионные градиенты. Этот комплекс процессов, объединяемых под общим названием «механизм электрической возбудимости», является яркой функциональной характеристикой нервной клетки. Возможность направленного распространения нервного импульса обеспечивается наличием у нервной клетки ветвящихся отростков, нередко простирающихся на значительные расстояния от её сомы и обладающих в области своих окончаний механизмом передачи сигнала через межклеточную щель на последующие клетки. Применение микроэлектродной техники позволило выполнить тонкие измерения, характеризующие основные электрофизиологические характеристики нервных клеток [Костюк, Крышталь, 1981; Окс, 1974; Ходоров, 1974]. Измерения показали, что каждая нервная клетка имеет отрицательный заряд, величина которого равна -40 – -65 мВ. Главное отличие нервной клетки от любой другой заключается в том, что она способна быстро изменять величину заряда вплоть до противоположного. Критический уровень деполяризации нейрона, при достижении которого возникает быстрый разряд, называется порогом генерации потенциала действия (ПД). Длительность потенциала действия различна у позвоночных и беспозвоночных животных – у беспозвоночных она равна 0,1 мс, а у беспозвоночных 1–2 мс. Серия потенциалов действия, распределённых во времени, является основой для пространственно-временного кодирования. Внешняя мембрана нейронов чувствительна к действию специальных веществ, которые выделяются из пресинаптической терминали – к нейромедиаторам. В настоящее время идентифицировано около 100 веществ, которые выполняют эту функцию. На внешней стороне мембраны расположены специализированные белковые молекулы – рецепторы, которые и взаимодействуют с нейромедиатором. В результате происходит открытие каналов специфической ионной проницаемости – только определённые ионы могут массированно проходить в клетку после действия медиатора. Развивается локальная деполяризация или гиперполяризация мембраны, которая называется постсинаптическим потенциалом (ПСП). ПСП могут быть возбудительными (ВПСП) и тормозными (ТПСП). Амплитуда ПСП может достигать 20 мВ.
Пейсмекер
Один из удивительных видов электрической активности нейронов, регистрируемой внутриклеточным микроэлектродом, – это пейсмекерные потенциалы. А. Арванитаки и Н. Халазонитис [Arvanitaki, Chalazonitis, 1955] впервые описали осциллирующие потенциалы нервной клетки, не связанные с поступлением к ней синаптических влияний. Эти колебания в ряде случаев могут приобретать такой размах, что превышают критический уровень потенциала, необходимый для активации механизма электрической возбудимости. Наличие в соме клетки таких волн мембранного потенциала было обнаружено на нейронах моллюсков. Они были расценены как проявление спонтанной, или ауторитмической активности, имеющей эндогенное происхождение [Alving, 1968]. Аналогичные ритмические колебания были затем описаны и во многих других типах нейронов. Способность к длительной ритмической активности сохраняется у некоторых клеток в течение длительного времени после полного их выделения [Chen et al., 1971; Греченко, Соколов, 1986]. Следовательно, в её основе действительно лежат эндогенные процессы, приводящие к периодическому изменению ионной проницаемости поверхностной мембраны. Важную роль играют изменения ионной проницаемости мембраны под действием некоторых цитоплазматических факторов, например системы обмена циклических нуклеотидов. Изменения активности этой системы при действии на соматическую мембрану некоторых гормонов или других внесинаптических химических влияний могут модулировать ритмическую активность клетки (эндогенная модуляция). Запускать генерацию колебаний мембранного потенциала могут синаптические и внесинаптические влияния. Л. Тауц и Г.М. Гершенфельд [Tauc, Gerschenfeld, 1960] обнаружили, что соматическая мембрана нейронов моллюсков, не имеющая на своей поверхности синаптических окончаний, обладает высокой чувствительностью к медиаторным веществам и, следовательно, имеет молекулярные хемоуправляемые структуры, свойственные постсинаптической мембране. Наличие внесинаптической рецепции показывает возможность модуляции пейсмекерной активности диффузным действием выделяющихся медиаторных веществ. Сложившаяся концепция о двух типах мембранных структур – электровозбудимой и электроневозбудимой, но химически возбудимой, заложила основу представлений о нейроне как пороговом устройстве, обладающем свойством суммации возбуждающих и тормозных синаптических потенциалов. Принципиально новое, что вносит эндогенный пейсмекерный потенциал в функционирование нейрона, заключается в следующем: пейсмекерный потенциал превращает нейрон из сумматора синаптических потенциалов в генератор. Представление о нейроне как управляемом генераторе заставляет по-новому взглянуть на организацию многих функций нейрона. Пейсмекерными потенциалами в собственном смысле этого слова называют близкие к синусоидальным колебания частотой 0,1–10 Гц и амплитудой 5–10 мВ. Именно эта категория эндогенных потенциалов, связанных с активным транспортом ионов, образует механизм внутреннего генератора нейрона, обеспечивающего периодическое достижение порога генерации ПД в отсутствие внешнего источника возбуждения. В самом общем виде нейрон состоит из электровозбудимой мембраны, химически возбудимой мембраны и локуса генерации пейсмекерной активности. Именно пейсмекерный потенциал, взаимодействующий с хемовозбудимой и электровозбудимой мембраной, делает нейрон устройством со «встроенным» управляемым генератором [Bullock, 1984]. Если локальный потенциал является частным случаем механизма генерации ПД, то пейсмекерный потенциал принадлежит к особому классу потенциалов – электрогенному эффекту активного транспорта ионов. Особенности ионных механизмов электрической возбудимости соматической мембраны лежат в основе важных свойств нервной клетки, в первую очередь её способности генерировать ритмические разряды нервных импульсов. Электрогенный эффект активного транспорта возникает в результате несбалансированного переноса ионов в разных направлениях. Широко известен гиперполяризационный постоянный потенциал как результат активного вывода ионов натрия, суммирующийся потенциалом Нернста [Ходоров, 1974]. Дополнительное включение активного насоса ионов натрия создаёт фазичекие медленные волны гиперполяризации (негативные отклонения от уровня мембранного потенциала покоя), обычно возникающие вслед за высокочастотной группой ПД, которая приводит к избыточному накоплению натрия в нейроне. Несомненно, что некоторые из компонентов механизма электрической возбудимости соматической мембраны, а именно электроуправляемые кальциевые каналы, вместе с тем являются фактором, сопрягающим мембранную активность с цитоплазматическими процессами, в частности с процессами протоплазматического транспорта и нервной трофики. Детальное выяснение этого важного вопроса требует дальнейшего экспериментального изучения. Пейсмекерный механизм, являясь эндогенным по происхождению, может активироваться и инактивироваться на длительное время в результате афферентных воздействий на нейрон. Пластические реакции нейрона могут обеспечиваться изменениями эффективности синаптической передачи и возбудимости пейсмекерного механизма (Соколов, Тавкхелидзе, 1975). Пейсмекерный потенциал является компактным способом передачи внутринейронной генетической информации. Приводя к генерации ПД, он обеспечивает возможность выхода эндогенных сигналов на другие нейроны, в том числе и эффекторные, обеспечивающие реакцию [Bullock, 1984]. Тот факт, что генетическая программа включает звено управления пейсмекерным потенциалом, позволяет нейрону реализовывать последовательность своих генетических программ. Наконец пейсмекерный потенциал в той или иной степени может подвергаться синаптическим влияниям. Этот путь позволяет интегрировать генетические программы с текущей активностью, обеспечивая гибкое управление последовательными программами. Пластические изменения пейсмекерного потенциала ещё больше расширяют возможность приспособления наследственно фиксированных форм к потребностям организма. Пластические изменения развиваются в этом случае не в геноме, а на пути выхода наследственной программы на реализацию (на уровне генерации ПД).
|
||
|
|
Последнее изменение этой страницы: 2018-04-12; просмотров: 529. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |