|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
РЕФЛЕКТОРНЫЙ УРОВЕНЬ ОРГАНИЗАЦИИ ДВИЖЕНИЙСпинной мозг — самый каудальный отдел ЦНС. Он состоит из 36-37 сегментов (рис. 5.1). От каждого сегмента спинного мозга отходят две пары (передних, или вентральных и дорсальных, или задних) нервов. Всего насчитывается 36-37 пар спинномозговых нервов: 8 пар шейных, 13 пар грудных, 7 пар поясничных, 3 пары крестцовых и 5—6 хвостовых. В верхних отделах спинного мозга есть полное соответствие между сегментами и позвонками, а в нижних этого соответствия нет. Поэтому нервы в виде пучка (так называемого конского хвоста) спускаются вниз к соответствующим межпозвоночным отверстиям и затем покидают позвоночный столб. У человека в связи с развитием верхних конечностей (рук) и нижних конечностей (ног) в спинном мозге образуются два утоления: шейное (ограничено нижними шейными и первым грудным сегментами спинного мозга) и поясничное (ограничено поясничными сегментами). Оба утолщения соответствуют областям отхождения толстых нервов, иннервирующих верхние и нижние пояса конечностей. Передние и задние корешки после выхода из спинного мозга вначале идут раздельно. У входа в костную щель позвоночного столба на дорсальном корешке находится утолщение (межпозвоночный ганглий), образованное афферентными нервными клетками, отростки которых и составляют дорсальный корешок. После выхода
Рис. 5.1. Спинной мозг человека.
из костного канала оба корешка сливаются и образуют смешанный кожно-мышечный нерв. В области поясничного и шейного утолщений корешки от нескольких сегментов после выхода из позвоночного канала переплетаются между собой, образуя поясничное и плечевое сплетения соответственно.  Нервы, иннервирующие мышцы, сухожилия и суставы, называются соматическими; иннервирующие внутренние органы, гладкие мышцы, сосуды, железы и оболочки — висцеральными. Эти два типа нервов имеют в своем составе как афферентные, так и эфферентные волокна. Серое и белое вещество спинного мозга. На поперечном сечении спинного мозга центральное серое вещество, образованное нервными клетками и их дендритами, напоминает форму бабочки (рис. 5.2). По периферии серого вещества располагается белое вещество, которое образовано преимущественно миелинизированными волокнами моторных (нисходящих) и сенсорных (восходящих) путей спинного мозга.
Рис. 5.2. Строение поясничного сегмента спинного мозга (схема). Слева — схема строения серого вещества, справа — основные проводящие пути в белом веществе. 1 — передний пирамидальный тракт; 2 — комиссуральное ядро; 3 — моторное ядро; 4 — промежуточное ядро; 5 — собственное ядро спинного мозга; 6 — желатинозная субстанция; 7 — пирамидный тракт; 8 — руброспинальный тракт; 9 — дорсальный спиномозжечковый тракт; 10 — (вентральный спиномозжечковый тракт; 11 — латеральный спиноталамический тракт; 12 — передний спиноталамический тракт; 13 — ретикулоспинальный тракт; 14 — латеральный вестибулоспиналъный тракт; 15 — дорсальные столбы.
В сером веществе различают передние и задние рога. В задних рогах выделяют краевую зону, желатинозную субстанцию и основание рога. В грудном отделе серое вещество образует дополнительно боковые рога, в которых находятся нейроны вегетативной нервной системы. Серое вещество спинного мозга содержит нервные клетки разных типов, разбросанные диффузно или собранные в виде ядер (показаны на левой половине рис. 5.2). Большинство ядер в спинном мозге занимают несколько сегментов, поэтому афферентные и эфферентные волокна, связанные с ними, входят в спинной мозг и выходят из него по нескольким корешкам. Наиболее значительными спинномозговыми ядрами являются ядра передних рогов, образованные мотонейронами. Выделяют три типа мотонейронов: альфа, бета и гамма. Альфа-мотонейроны представлены крупными мультиполярными клетками с диаметром тела 25-75 мкм; их аксоны иннервируют двигательные мышцы, которые способны развивать значительную силу. Бета-мотонейроны — это мелкие нейроны, которые иннервируют тонические мышцы. Гамма-мотонейроны еще мельче — диаметр их тела 15-25 мкм. Они локализованы в двигательных ядрах вентральных рогов среди альфа- и бета-мотонейронов. Гамма-мотонейроны осуществляют двигательную иннервацию мышечных рецепторов (мышечных веретен). Аксоны этих двух типов мотонейронов составляют основную массу передних корешков спинного мозга. В основании заднего рога находится другое скопление нервных клеток, образующих промежуточное ядро спинного мозга. Нейроны, составляющие ядро этой зоны, имеют короткие аксоны, которые в основном идут в передний рог и образуют там синаптические контакты с мотонейронами. Аксоны некоторых из этих нейронов распространяются на 2-3 сегмента, но никогда не выходят за пределы спинного мозга. По поверхности заднего рога располагается слой мелких нейронов, образующих желатинозную субстанцию (роландову), которые осуществляют контроль синаптического проведения от первичных афферентов на нейронные системы спинного мозга. Рефлексы спинного мозга. Выдающуюся роль в философском осмысливании механизмов деятельности нервной системы сыграли работы французского философа и математика Р. Декарта. Он впервые выдвинул принцип рефлекторной деятельности организма. Термин «рефлекс», которым сейчас широко пользуются, был предложен значительно позже чешским физиологом И. Прохазкой. Рефлексом называется ответная реакция организма на сенсорное воздействие. Рефлекс осуществляется через рефлекторную дугу (цепочку нейронов) нервной системы. Например, в ответ на стимуляцию кожи рука отдергивается. На языке нейрофизиологии это означает, что в ответ на стимуляцию возбуждаются афферентные волокна от кожи, далее это возбуждение через задние корешки спинного мозга достигает соответствующих мотонейронов (в передних рогах серого вещества спинного мозга), и по их аксонам двигательная команда достигает соответствующих мышц. Введем некоторые важные определения. Минимальная сила раздражения, которая вызывает данный рефлекс, называется порогом (или пороговым стимулом) данного рефлекса. Всякий рефлекс имеет рецептивное поле, т. е. совокупность рецепторов, раздражение которых вызывает рефлекс с наименьшим порогом. При изучении движений приходится расчленять сложный рефлекторный акт на отдельные относительно простые рефлексы. Вместе с тем следует помнить, что в естественных условиях отдельный рефлекс выступает только как элемент сложной деятельности. Простейшими рефлексами, которые можно легко наблюдать, являются сгибательный и разгибательный. Под сгибанием следует понимать уменьшение угла данного сустава, а под разгибанием — его увеличение. Сгибательные рефлексы широко представлены в движениях человека. Характерным для этих рефлексов является большая сила, которую они могут развивать. Вместе с тем они быстро утомляются. Разгибательные рефлексы также широко представлены в движениях человека. Например, к ним относятся рефлексы поддержания вертикальной позы. Эти рефлексы в отличие от сгибательных значительно более устойчивы к утомлению. Действительно, мы можем долго ходить и стоять, но для выполнения длительной работы, например поднимания рукой гири, наши физические возможности значительно более ограничены. Универсальный принцип рефлекторной деятельности спинного мозга получил название общего конечного пути. Дело в том, что соотношение количества волокон в афферентных (задних корешках) и эфферентных (передних корешках) путях спинного мозга составляет примерно 5:1. Ч. Шеррингтон (выдающийся английский физиолог, современник И. П. Павлова) образно сравнивал этот принцип с воронкой (воронка Шеррингтона), широкую часть которой составляют афферентные пути задних корешков, а узкую — эфферентные пути передних корешков спинного мозга. Нужно помнить, что общий конечный путь, являясь при осуществлении рефлекторной деятельности спинного мозга функциональным образованием, часто оказывается сложным. Нередко территория конечного пути одного рефлекса перекрывается с территорией конечного пути другого рефлекса. Другими словами, разные рефлексы могут соревноваться за занятие конечного пути. Это можно проиллюстрировать таким примером. Представим себе, что собака убегает от опасности и ее в это время кусает блоха. В данном примере за общий конечный путь — мышцы задней лапы — соревнуются два рефлекса: один — чесательный, а другой — рефлекс ходьбы-бега. В одни моменты может пересиливать чесательный рефлекс, и собака останавливается и начинает чесаться, но затем вновь верх может взять рефлекс ходьбы-бега, и собака возобновит бег. Как уже указывалось, при осуществлении рефлекторной деятельности отдельные рефлексы спинного мозга взаимодействуют
Рис. 5.3. Коленный рефлекс.
друг с другом, образуя функциональные системы. Один из самых важных элементов функциональной системы — обратная афферентация, благодаря которой нервные центры как бы оценивают, как выполняется реакция, и могут внести в нее необходимые коррективы. Некоторые конкретные механизмы обратной афферентации будут рассмотрены ниже. Один из таких примеров — рефлекс на растяжение. Примером такого рефлекса может служить коленный рефлекс, который возникает при легком ударе по сухожилию мышцы в подколенной чашечке (рис. 5.3). Рефлекс на растяжение предотвращает чрезмерное растяжение мышцы, которая как бы сопротивляется растягиванию. Этот рефлекс возникает как ответная реакция мышцы на стимуляцию ее рецепторов, поэтому его часто обозначают как собственный рефлекс мышцы. Известны два вида рефлекса на растяжение: тонический (медленный) и фазический (быстрый). Для разделения этих двух типов рефлексов применяют два способа растягивания мышцы: медленное растягивание мышцы вызывает тонический рефлекс на растяжение, быстрое — фазический рефлекс. Примерами фазических рефлексов на растяжение могут служить коленный и ахиллов (рефлекс вызывают легким ударом по ахиллову сухожилию). Доказательством того, что эти рефлексы осуществляются за счет активации рецепторов только самих мышц, а не сухожилий или суставов, может служить их сохранение при анестезии суставной сумки. Примером тонического рефлекса на растяжение может быть собственный рефлекс икроножной мышцы. Это одна из главных мышц, благодаря которой поддерживается вертикальная поза человека. Физиология опорно-двигательного аппарата. Познакомимся с некоторыми принципами деятельности мышц нашего тела. С функциональной точки зрения мышца состоит из двигательных единиц. Двигательная единица — это группа мышечных волокон, которые иннервируются одним мотонейроном и поэтому функционируют как единое целое по принципу «все или ничего» (возбуждается или не возбуждается). Среднее количество мышечных волокон, приходящихся на одну двигательную единицу, варьирует в разных мышцах от 4-6 волокон, участвующих в тонких движениях (например, глазодвигательная мышца), до 2000 — у больших мышц, находящихся в постоянном тоническом сокращении (например, икроножная мышца). Территория двигательной единицы на поперечном срезе близка к кругу, и, как правило, одну и ту же территорию занимают 2—3 двигательные единицы. Другими словами, имеет место перекрытие отдельных территорий, занимаемых двигательными единицами. В связи с тем что каждая двигательная единица сокращается по принципу «все или ничего», такая структура мышцы обеспечивает гладкость ее сокращения в условиях, когда отдельные двигательные единицы работают в режиме одиночных сокращений. Поэтому градация силы сокращения целой мышцы осуществляется преимущественно за счет вовлечения в деятельность новых двигательных единиц. В естественных условиях «гладкость» сокращения мышцы обеспечивается асинхронностью работы разных двигательных единиц. В мышцах человека, а также других млекопитающих есть два типа двигательных единиц — быстрые и медленные. Быстрые двигательные единицы иннервируются крупными мотонейронами и могут развивать большую силу, но быстро утомляются. В отличие
Рис. 5.4. Кривая длина-напряжение для тетануса медленной мышц (т. soleus) кошки.
Вертикальные линии на кружках указывают ошибку измерения. Стрелкой на оси абсцисс отмечена длина мышцы, при которой стопа образует прямой угол с голенью. от них медленные двигательные единицы иннервируются мелкими мотонейронами и могут длительно поддерживать активность тонического типа. Наличие двух типов двигательных единиц — быстрых фазических и медленных тонических — является приспособлением мышцы для работы в широком диапазоне силовых нагрузок. Мышцы млекопитающих животных хорошо приспособлены для выполнения своих функций. Известно, что максимальное напряжение при тетанусе мышца развивает только при своей «нормальной» длине, т. е. при той, которую она имеет в состоянии покоя в организме. Специальные исследования показали, что эти оптимальные условия для сокращения мышцы (развития максимальной силы) соответствуют и физиологическому диапазону движения в суставе. На рис. 5.4 приведена кривая длина-напряжение камбаловидной мышцы, полученная при изометрическом сокращении. Видно, что кривая тетанического напряжения мышцы имеет большое плато, охватывающее диапазон около 12 мм. Это имеет важное значение при длительных рефлексах позы: проекция плато на ось абсцисс примерно совпадает с отрезком, выделенным жирной линией, которая соответствует диапазону возможных положений голеностопного сустава у интактной кошки от максимально разогнутого до максимально согнутого. Существует зависимость между характеристиками двигательной единицы и характеристиками соответствующего мотонейрона. Так, низкопороговые маленькие (диаметр сомы до 20 мкм) мотонейроны с низкой скоростью проведения по аксонам в рефлекторных реакциях ведут себя как тонические и иннервируют медленные двигательные единицы, которые развивают небольшую силу, но устойчивы к утомлению и поэтому способны к длительной работе. Высокопороговые большие (диаметр сомы 50—70 мкм) мотонейроны с высокой скоростью проведения по аксону ведут себя в рефлекторных реакциях как фазические и иннервируют быстрые двигательные единицы. Сила, развиваемая быстрыми двигательными единицами, относительно велика, но они хуже приспособлены к длительной работе. Эти экспериментальные данные позволили высказать гипотезу, что в условиях диффузной активации мотонейронов, иннервирующих данную мышцу из вышележащих мозговых структур, организующая роль в возбуждении мышцы принадлежит порогам отдельных мотонейронов. Эти пороги определяются морфологическими размерами мотонейронов (принцип размера): маленькие нейроны имеют низкий, а большие — более высокий порог активации. Благодаря этому принципу обеспечивается автоматический выбор активируемых двигательных единиц в условиях той или иной двигательной задачи. Вначале включаются медленные тонические мышцы, а затем на их фоне сокращаются быстрые фазические мышцы. Кроме двигательных мышечных волокон, в мышце содержатся также многочисленные рецепторы. Они составляют две группы: мышечные рецепторы (так называемые мышечные веретена) и рецепторы сухожилий. Особенно большое количество мышечных веретен находится в мышцах, выполняющих тонкие движения. Например, в мышцах передних лап кошки находится 50-100 веретен на 1 г мышечной массы. Мышечные веретена имеют также моторное управление, благодаря которому их чувствительность может задаваться из спинного мозга. Положение мышечного веретена относительно мышечного волокна показано на рис. 5.5. Мышечное веретено в целой мышце закреплено параллельно отдельным двигательным мышечным волокнам. Мышечные веретена передают в нервные центры сигналы о длине мышцы и о скорости ее изменения. Обратите внимание, что мышечные веретена в мышце расположены параллельно основным мышечным волокнам. При сокращении мышцы мышечное веретено разгружается и перестает
Рис. 5.5. Схема взаимоотношений сокращения мышцы и реакции мышечного веретена. Слева — растяжение мышечного волокна с параллельно закрепленным мышечным веретеном (А): справа — механограммы растяжения мышечного волокна и импульсации в рецепторе мышечного веретена. При укорочении (сокращении) мышечного волокна происходит разгрузка мышечного веретена (Б, слева) и прекращение импульсации в соответствующем рецепторе. Для предотвращения разгрузки мышечного веретена активируется гамма-мотонейрон (В). Благодаря этому спинной мозг продолжает получать информацию о длине мышцы; 1 — мышечное волокно; 2 — мышечное веретено; 3 — стимуляция альфа-мотонейрона; 4 — стимуляция гамма-мотонейрона.
сообщать в ЦНС о длине и изменении длины мышцы. Чтобы этого не происходило, мышечные веретена имеют специальное моторное управление — они могут активироваться через аксоны гамма-мотонейронов. В отличие от этого сухожильный рецептор находится в сухожилии, которым мышца крепится к кости. Поэтому рецептор тем больше возбуждается, чем большая сила растягивает сухожилие. Другими словами, сухожильный рецептор измеряет силу, с которой мышца тянет сухожилие. Таким образом, состояние мышцы контролируется двумя типами рецепторов — мышечными веретенами, передающими информацию о длине мышцы и скорости изменения этой длины, и сухожильным рецептором, который сигнализирует о силе мышечного сокращения. Мотонейроны передних рогов серого вещества спинного мозга отдают аксоны, которые иннервируют двигательные мышцы. В пределах серого вещества эти аксоны отдают возвратные коллатерали (ответвления), которые заканчиваются на специальных клетках (клетках Реншоу); эти последние, в свою очередь, заканчиваются на мотонейронах тормозными синапсами. Таким образом, мотонейроны находятся под влиянием отрицательной обратной связи. Благодаря этой связи частота мотонейрона обычно бывает в пределах 10-15 имп/с. Сила, развиваемая мышцей, определяется количеством включенных в деятельность двигательных единиц. В работающей мышце активные двигательные единицы постоянно меняются, и этим достигается плавность сокращения всей мышцы и увеличение ее работоспособности. Что будет, если отключить тормозные влияния на мотонейроны через клетки Реншоу? Известно, например, что столбнячный токсин блокирует передачу тормозного влияния с клетки Реншоу на мотонейрон. Основным признаком столбняка являются судороги, тетанус мышц. Можно сделать вывод, что отключение этой обратной связи приводит к неконтролируемому возбуждению мотонейрона и несанкционированному сокращению (тетанусу) мышц. Таким образом, благодаря активности клеток Реншоу импульсация мотонейронов поддерживается в оптимальном диапазоне, необходимом для управляемого сокращения мышцы. Сухожильные рецепторы (измерители силы) оказывают на мотонейроны тормозное действие. В качестве примера рассмотрим «рефлекс складного ножа», описанный еще в начале XX века выдающимся английским физиологом Ч. Шеррингтоном. Этот рефлекс проявляется следующим образом. При растяжении мышцы включается, как было описано выше, рефлекс на растяжение, причем мышца сопротивляется этому растяжению. Однако если продолжить растягивать мышцу, она неожиданно расслабляется (вспомните, как трудно бывает открыть лезвие складного ножа, отсюда и название рефлекса). Специальные исследования показали, что в этом случае, т.е. при преодолении сопротивления мышцы,
Рис. 5.6. Поддержание вертикальной позы человеком. Слева — проекция центра тяжести (точка) на площадь опоры; а — вертикальная составляющая; б — горизонтальная составляющая колебания проекций центра тяжести.
происходит возбуждение сухожильного рецептора, вследствие чего тормозятся мотонейроны и вызывается неожиданное расслабление мышцы. Поза. Движения, которые совершает человек, характеризуются общим свойством — они всегда происходят из определенной позы. Смена поз осуществляется с помощью движений. Ниже будут более подробно рассмотрены отдельные рефлексы установки тела, головы и глаз в пространстве, которые нужно расценивать как частные случаи отдельных поз. Типичным примером позы является стояние человека. Он сохраняет равновесие благодаря тому, что центр тяжести находится в пределах площади опоры (рис. 5.6). Вертикальная ось тела проходит несколько впереди голеностопных и коленных суставов и несколько позади тазобедренных. Поддержание вертикальной позы является активным процессом, на который влияют не только внешние возмущения, но и возмущения, связанные, например, с дыханием. Общие принципы управления движениями. Управление движениями осуществляется ЦНС. Двигательная функция человека достигла наивысшего развития в связи с прямохождением и трудовой деятельностью. Поэтому в управление этой функции включены также высшие центры, включая кору больших полушарий. Это прежде всего относится к произвольным движениям — спортивным и рабочим движениям, речи и т. д. Очень важным биомеханическим свойством опорно-двигательного аппарата организма животных и человека является наличие в
Рис. 5.7. Ходьба (объяснение в тексте).
скелете большого числа степеней свободы. Это происходит вследствие многозвенности скелета, а также двух- и трехосности многих суставов. С одной стороны, многозвенность скелета обеспечивает большую свободу движений, но с другой — сильно затрудняет управление такой сложной системой. В каждом конкретном случае используются только некоторые движения, но ЦНС постоянно контролирует (ограничивает) остальные, что сообщает движению устойчивость. Нарушение этой функции ЦНС приводит к характерным патологическим изменениям движений, примеры которых будут приведены ниже (мозжечковые нарушения, нарушения, связанные с патологией стриатума, и пр.). Существуют два типа коррекции движений: при помощи обратных связей и на основе программы. Первый тип коррекций, как правило, выполняется при медленных движениях, тогда как второй — при быстрых. Примерами быстрых движений могут служить печатание на машинке, кидание мяча, движения глаз в орбитах и др. В коррекции движений активно участвуют не только мышечные, суставные, сухожильные и другие рецепторы такого типа, но также зрение, слух, вестибулярный аппарат. Участие мышц в конкретном движении весьма многообразно. Однако в функциональном отношении в конкретном двигательном акте выделяют основную мышцу (основной двигатель), вспомогательные мышцы и стабилизаторы (мышцы, фиксирующие суставы, но напрямую не участвующие в движении). Рассмотрим реализацию рассмотренных общих принципов на примере ходьбы. Ходьба является обычной формой локомоции человека и относится к циклическим двигательным актам. Выделяют следующие периоды ходьбы (рис. 5.7). Двухопорный период (1), когда обе ноги стоят на опоре: одноопорный период — для правой ноги (2) и переносный (3) — для левой. Снова двухопорный (4) и затем одноопорный (5) для левой ноги. Во время опорной фазы происходит перекатывание стопы с пятки на носок. В опорных стадиях выделяют передний и задний толчки: первый — при переносе нагрузки на опорную ногу, второй — при отталкивании ноги от опоры. В ходьбе участвуют мышцы стопы, голени, бедра и тазового пояса. Нейрофизиологические механизмы локомоции. Главным элементом нейрофизиологического механизма являются центральные генераторы движений. У позвоночных животных они находятся в спинном мозге и задают характер сокращения мышц конечности, поясов конечностей или соответствующих сегментов тела. Центральный генератор — это функциональное понятие. У позвоночных животных в состав центрального генератора входят нейроны нескольких сегментов спинного мозга; эффекторными нейронами являются мотонейроны. Под активацией генератора понимают перевод его из состояния покоя в состояние активности. Это осуществляется системой командных нейронов, расположенных на разных уровнях нервной системы. Активирующие сигналы представляют собой тонический поток импульсов, интенсивность которого и определяет уровень активности генератора. В основе координации движений конечностей и частей тела при локомоции лежит взаимодействие различных центральных генераторов. Это взаимодействие осуществляется специальными координирующими нейронами. Несмотря на то что центральный генератор может работать автономно, у интактного животного он подвержен мощному влиянию периферических афферентов. Благодаря этому в реальных условиях работа генератора адаптирует локомоцию к меняющимся условиям. Кроме этого, супраспинальные центры также оказывают воздействие на работу центрального генератора. Благодаря такому устройству система управления локомоцией у животных сочетает в себе принцип программного управления с управлением по принципу обратной связи, при этом обратные связи охватывают все иерархические уровни. Сеть короткоаксонных интернейронов, которые, как предполагают, и составляют локомоторный генератор, локализована в латеральных участках серого вещества спинного мозга. Благодаря активности короткоаксонных нейронов отдельные сегменты спинального генератора могут объединяться и работать как целое. Генератор одной конечности состоит из двух полуцентров (сгибательного и разгибательного), которые взаимодействуют реципрокно. Возможно, автоматизм работы локомоторного центра обеспечивается нейронами с пейсмекерными свойствами (способны к самовозбуждению). Исследования на животных показали, что между конечностями одного пояса (гомологичными) и обеими конечностями одной стороны тела (гомолатеральными) при разных походках реализуются только две программы взаимодействия: противофазная и синфазная. Переход от одного типа взаимодействия к другому происходит на протяжении 1—2 локомоторных циклов. Различные комбинации программ, по-видимому, лежат в основе различных типов походок. Некоторые экспериментальные наблюдения на животных дают основание полагать, что при локомоции генераторы задних конечностей доминируют, им присуща большая степень автоматизма; в отсутствие локомоции они тонически тормозят центры передних конечностей. У всех позвоночных животных (от рыб до приматов) обнаружены области мозга, электрическая стимуляция которых вызывает локомоцию. Усиление стимуляции локомоторной области сопровождается более энергичной локомоцией. Установлено, что для успешной локомоции необходима сохранность латеральной и вентролатеральной областей спинного мозга. У кошки в головном мозге описаны три локомоторные области: мезэнцефалическая, гипоталамическая и локомоторная полоски. Мезэнцефалическая локомоторная область занимает довольно протяженную область в дорсальном отделе покрышки среднего мозга. Гипоталамическая локомоторная область расположена билатерально над мамиллярными телами гипоталамуса. Локомоторная полоска простирается от мезэнцефалической локомоторной области, далее проходит через мост (вентролатеральнее двигательного ядра тройничного нерва) и продолговатый мозг (вентролатеральнее спинального ядра тройничного нерва). Далее эта полоска спускается в спинной мозг, и по ней происходит полисинаптическое распространение возбуждения в каудальном направлении. По мнению исследователей, мезэнцефалическая полоска представляет собой интегративный центр, находящийся между ростральными отделами мозгового ствола и спинным мозгом. Рефлексы установки глаз, головы и тела в пространстве. Раздражителями лабиринтных рецепторов являются угловые и линейные ускорения. Угловые ускорения воспринимаются полукружными каналами, а линейные — отолитовыми органами (мешочками). Импульсация, поступающая от вестибулярных рецепторов в ЦНС, вызывает вестибулярные рефлексы шеи, туловища, конечностей и глаз. Вестибулярный аппарат внутреннего уха вместе с вестибулярными ядрами мозгового ствола обеспечивает рефлекторный контроль положения туловища, головы и глаз. Анатомической основой этого контроля являются следующие системы: латеральный вестибулоспинальный путь, начинающийся от латерального вестибулярного ядра (Дейтерса); медиальный вестибулоспинальный путь, берущий начало от медиального вестибулярного ядра; ретикулоспинальные пути и путь от верхнего вестибулярного ядра (Бехтерева) на глазодвигательные мышцы. Рефлексы, обеспечивающие сохранение равновесия и положение тела в пространстве при спокойном лежании, стоянии и сидении в различных положениях, называются статическими рефлексами (рис. 5.8). Рефлексы, при помощи которых животное или человек совершает активные и пассивные движения и которые частично компенсируют последствия этих перемещений, называются статокинетическими рефлексами. Статические рефлексы, в свою очередь, делятся на две большие группы. В покое животное при помощи позных рефлексов принимает позу, т.е. определенное положение отдельных участков тела по отношению друг к другу, которое сопровождается закономерным распределением тонуса всей мускулатуры и тонической фиксацией в различных суставах. Те рефлексы, благодаря которым животное может стоять, называются рефлексами стояния. Вторую группу составляют рефлексы, благодаря которым животное может принимать нормальное положение из неестественного положения (например, вставать, правильно устанавливать голову и пр.). Эти рефлексы называют установочными. К группе статических рефлексов относят также компенсаторные движения глаз. Строение вестибулярного аппарата. У млекопитающих внутреннее ухо представляет трехцелевой орган: полукружные каналы служат для рецепции углового ускорения, в мешочках (утрикулюсе и саккулюсе) находятся отолитовые органы для регистрации линейного ускорения, а улитка с кортиевым органом является органом частотного анализа звука (рис. 5.9). Три полукружных канала (комплекс одной стороны черепа) расположены в плоскостях, образующих приближенно прямоугольную систему координат. Каждый канал выходит из утрикулюса и,
Рис. 5.8. Установочные рефлексы. А — шейные к лабиринтные рефлексы противодействуют друг другу; Б, Д, 3 — действуют одни шейные рефлексы; В — шейные и лабиринтные рефлексы усиливают друг друга; Г, Д, Ε — действуют одни лабиринтные рефлексы; Ж — шейные и лабиринтные рефлексы усиливают друг друга; И — шейные и лабиринтные рефлексы противодействуют друг другу.
описав дугу, близкую к полуокружности, опять приходит к нему, но с другой стороны. В непосредственной близости к утрикулюсу канал резко расширяется (примерно в 10 раз), образуя ампулу. В ампуле находится нейроэпителий, состоящий из сенсорных клеток, волосковидные отростки которых пронизывают канальцы (диаметром 3—5 мкм) студневидного колпачка — купулы (рис. 5.10).
Рис. 5.9. Подразделения вестибулярного нерва и расположение лабиринта в черепе человека. 1, 2, 3 — горизонтальный, задний, передний полукружные каналы; 4 — утрикулюс; 5 — саккулюс; 6, 7 — верхний и нижний ганглии Скарпа; 8 — вестибулярный нерв; 9 — лицевой нерв; 10 — улитковый нерв.
Отолитовый аппарат, представленный мешочками — саккулюсом и утрикулюсом, служит для измерения линейных ускорений, а с помощью полукружных каналов измеряют угловые ускорения. Принцип измерения в обоих органах одинаковый — движение более плотных частиц в жидкости меньшей плотности (рис. 5.10, 5.11). Большие отолиты (от 10 до 20 мм) состоят из известняка плотностью 2,93 (от плотности воды), имеющего различную консистенцию — от массы, подобной зубной пасте, до камней с характерной пластинчатой структурой. Отоконии у млекопитающих, включая человека, состоят из кальцита с плотностью 2,71 (размеры от 1 до 20 мкм). Протеиновая сеть, в которую включены отоконии, движется как единое целое. Отолитовая мембрана пронизана волосковыми отростками (цилиями) сенсорных клеток, которые вместе с поддерживающими клетками образуют сенсорный эпителий — макулу. Между отолитами и макулой находится пространство
Рис. 5.10. Схема строения отолитового органа (продольный разрез отолитовой мембраны и макулы). 1 — отолитовая мембрана; 2 — рецепторы; 3 - нерв; 4 — субмембранное пространство; 5 — поддерживающие клетки.
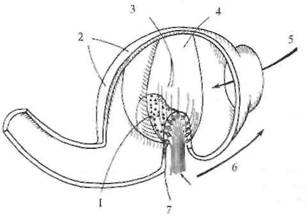
Рис. 5. 11. Схема строения ампулярного органа. Цилии волосковых клеток (показано только по одной цилии на рецептор) входят в купулу, которая движется вместе с эндолимфой при движении головы. 1 — crista ampullaris; 2— ампула; 3 — цилии; 4 — купула; 5 — эндолимфа; 6 — направление движения головы; 7 — слой рецепторов.
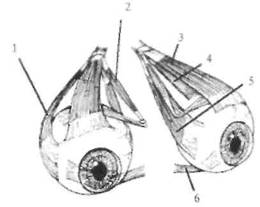
заполненное желеобразной массой. Благодаря этому отолит скользит по макуле и деформирует волоски чувствительных клеток (см. рис, 5.11). Максимальное смещение отолита по макуле для саккулюса составляет 0,1 мм, для утрикулюса — 0,005 мм. Нейроэпителий утрикулюса лежит горизонтально, а саккуляр-ный — парасагиттально. В результате этого утрикулярные рецепторы чувствительны к небольшим наклонам головы от ее нормального положения и к линейным ускорениям, которые возникают во время перемещения в горизонтальной плоскости. Саккулярные единицы в отличие от этого чувствительны к дорсовентральному ускорению, как это имеет место при прыжках и падениях. В противоположность широким вариациям размеров тела у разных животных размеры полукружных каналов примерно одинаковы, также как размеры компаса мало зависят от размера корабля. Волосковые клетки в нейроэпителий как отолитовых органов (саккулюсе и утрикулюсе), так и в ампулах полукружных каналов расположены упорядоченными рядами. Благодаря этому, например, отклонение купулы в сторону канала в горизонтальных каналах тормозит спонтанный разряд рецепторов, а в обратную сторону — увеличивает. В вертикальных каналах ориентация волосковых рецепторов противоположна." отклонение в сторону утрикулюса является тормозным, а в сторону канала — возбуждающим. В физиологических пределах усиление импульсации кристы при ускорении изменяется как линейная функция этого ускорения. Таким образом, каждый полукружный канал регистрирует степень ускорения в своей плоскости. В связи с тем что все три канала практически ортогональны (перпендикулярны относительно друг друга), они передают в ЦНС точную информацию об амплитуде и направлении углового ускорения головы в пространстве. Краткие сведения об устройстве периферического звена глазодвигательной системы. Глазное яблоко управляется тремя парами мышц: двумя парами прямых и двумя косыми (медиальной и латеральной) (рис. 5,12). Каждая пара работает по принципу «тяни-толкай». Как уже указывалось, на каждой стороне черепа имеются три полукружных канала и отолитовые органы, представленные утрикулюсом и саккулюсом. Полукружные каналы одной стороны образуют прямоугольную систему координат. Каналы противоположных сторон, лежащие в одной плоскости, взаимодействуют между собой по принципу «тяни-толкай»: при вращении головы один канал пары максимально возбуждается, а другой максимально
рис. 5.12. Окуломоторные мышцы. 1 — наружная прямая мышца; 2 — верхняя косая мышца; 3 — верхняя прямая мышца; 4 — нижняя прямая мышца; 5 — внутренняя прямая мышца; 6 — нижняя косая мышца.
тормозится. Таким образом, все полукружные каналы образуют три пары: правый — левый горизонтальные, левый передний — правый задний и левый задний — правый передний. Каждая пара каналов связана с определенной парой глазодвигательных мышц трехнейронной цепочкой: горизонтальные каналы проецируются к горизонтальным прямым мышцам, левый передний — правый задний — к левой вертикальной прямой и правой косой, правый передний — левый задний каналы — к правой вертикальной и левой косой. Установочные рефлексы мозгового ствола. Даже таламическое животное (т.е. животное, у которого удалены большие полушария) способно принимать и поддерживать нормальное положение тела. Это обеспечивается совокупным действием ряда рефлексов мозгового ствола. Примеры этих рефлексов приведены ниже. 1. Лабиринтный установочный рефлекс на голову состоит в том, что при любом положении тела голова животного принимает нормальное положение с параллельно расположенной ротовой щелью благодаря рефлексу с вестибулярного аппарата. 2. Тонические лабиринтные рефлексы на глаза. Компенсаторные движения глаз при поворотах головы и туловища можно наблюдать у всех позвоночных. Однако у животных с боковым положением глаз (например, у кролика) они выражены более отчетливо. Под действием этих рефлексов глаз максимально перемешается назад, когда голова животного занимает положение мордой вниз. Максимальный поворот глаза верхним полюсом роговицы вверх происходит при положении головы мордой вертикально вверх. Благодаря этим рефлексам каждому положению головы в пространстве соответствует определенное положение глазного яблока в орбите. При этом не имеет значения, с какой стороны достигалось положение головы. Всегда устанавливается одно и то же закономерное положение глаз. У животных с фронтальным расположением глаз (например, у хищных, приматов) при наклоне головы вперед глаза отклоняются вверх. Тонические реакции глаз рефлекторно возникают в результате раздражения отолитового аппарата. При повороте головы кролика вокруг сагиттальной плоскости также возникают тонические лабиринтные рефлексы на глаза. При этом отклонения обоих глаз при разных положениях головы в пространстве происходят в противоположных направлениях. Если вращать голову от нормального положения (ротовая щель расположена горизонтально), то глаз, находящийся внизу, отклонится в орбите вверх. Отклонение достигает максимума при положении головы на боку. Контралатеральный глаз при этом отклонится вниз. При вращении через положение на затылке реакция глаз быстро изменяется на обратную. Таким образом, существуют два независимых компонента тонического лабиринтного рефлекса на глаза, которые образуют все возможные комбинации при каждом положении головы в пространстве. Быстрые движения глаз, поскольку они вызываются с полукружных каналов, не участвуют в этих статических рефлексах. Активирование полукружных каналов (вращением головы) вызывает вестибулоокулярный рефлекс. 3. Вестибулоокулярный рефлекс (ВОР) — один из древнейших; он сопровождает позвоночных животных на протяжении всей их эволюции. Рефлекс проявляется в том, что при вращении головы глаза в орбитах противовращаются. Внешне это проявляется в виде нистагма — колебательных движений глазных яблок в орбитах. Различают медленную компоненту нистагма, при которой глаза вращаются в сторону вращения головы примерно с той же скоростью, и быструю фазу нистагма, при которой глаза в орбитах после достижения своего крайнего положения скачком (саккадой) возвращаются в исходное положение. Этот рефлекс можно наблюдать
Рис. 5.13. Схема нейронных цепей, обеспечивающих пластичность вестибулоокулярного рефлекса с участием флоккулюса мозжечка. 1 — глаз; 2 — нижняя олива; 3 — клетка Пуркинье коры мозжечка; 4 — клетка-зерно коры мозжечка; 5 — полукружный канал; 6 — мшистое волокно; 7 — мотонейрон; 8 — вестибулярное ядро.
как в полной темноте, так и на свету. Биологическая роль этого рефлекса — предотвращать сползание сетчаточного изображения при вращении головы. Входным сигналом для ВОР является импульсация, передаваемая по вестибулярным афферентам от полукружных каналов. Вестибулярные афференты направляются в мозговой ствол (верхнее вестибулярное ядро). Этот афферентный сигнал после обработки выходит на эффекторное звено — экстраокулярные мышцы. Главная проблема для такой системы состоит в калибровке: как отрегулировать скорость противовращения глазных яблок, чтобы оно эффективно компенсировало вращение головы. На рис. 5.13 показана принципиальная схема ВОР. В процессе калибровки скорости противовращения глазных яблок в орбитах относительно направления вращения головы участвует мозжечок. Это символически показано одной клеткой коры мозжечка (клеткой Пуркинье), а именно — самой задней частью мозжечка (флоккулонодулярная), которую часто называют вестибулярным мозжечком (см. приложение 6). Гипотеза об участии флоккулюса мозжечка в пластичности ВОР была высказана на основе тесных связей вестибулярного мозжечка (flocculus и nodulus) как с вестибулярной, так и со зрительной системой. В пользу этого предположения говорят также результаты по удалению вестибулярного мозжечка. Оказалось, после удаления вестибулярного мозжечка (флоккулюса и парафлоккулюса), а также нижней половины VIII дольки и нижних 3-4 долек парамедианной доли усиление ВОР устанавливалось на уровне 1,063 (диапазон 0,95-1,173) и исчезала способность БОР к пластическим изменениям коэффициента усиления. Предполагают, что сигналом для изменения усиления ВОР является смещение сетчаточного изображения. Далее этот сигнал по дополнительной зрительной системе поступает в претектальную область (ядро оптического тракта), затем к нижней оливе и через лазающие волокна из нижней оливы к клеткам Пуркинье коры вестибулярного мозжечка (флоккулюсу). Сигнал скорости вращения головы передается от верхнего вестибулярного ядра по мшистым волокнам к клеткам-зернам и далее через параллельные волокна поступает также на клетки Пуркинье. Таким образом, клетки Пуркинье коры вестибулярного мозжечка выполняют роль компараторов (сравнителей), благодаря «работе» которых может модифицироваться коэффициент усиления ВОР. У человека ВОР участвует в реакции установки взора. Эта реакция возникает при появлении сбоку в поле зрения неожиданного зрительного стимула. В первый момент человек скачкообразно переводит глаза в сторону стимула и «захватывает» зрением этот стимул. Слово «захватывает» означает, что стимул помещается в область сетчатки с наилучшим разрешением (зрительная ямка, fovea). С небольшим отставанием (это связано с инерцией) в эту же сторону начинает перемешаться голова. Чтобы изображение не сползло с сетчатки (другими словами, чтобы человек не потерял изображение), включается ВОР, который в данном случае запускается благодаря вращению головы и, следовательно, возбуждению рецепторов полукружных каналов. При этом скорость противовращения глаз в орбитах равна скорости вращения головы. Благодаря этому взор (алгебраическая сумма скоростей вращения глаз и головы) остается неподвижным в пространстве. ВОР обеспечивает движение глаза в противофазе с движениями головы в диапазоне 0,01—6,5 Гц. На более низкой частоте в стабилизации сетчаточного изображения участвуют также оптомоторный рефлекс и рефлекс прослеживания. Пол «стабилизацией» будем понимать то, что положение глаза относительно зрительной цели не меняется, несмотря на вращение головы. Это достигается механизмом ВОР: глаза противовращаются в орбитах со скоростью, равной скорости вращения головы. Для успешной стабилизации сетчаточного изображения при вращениях головы необходимо регулировать коэффициент усиления ВОР. Нетрудно себе представить ситуации, в которых необходимо изменять усиление ВОР. Например, при росте организма меняются размеры головы и глазных яблок, в силу разных обстоятельств (травмы, процессы старения и пр.) меняется упругость глазодвигательных мышц и многое другое. Коэффициент усиления ВОР регулируется в коре задней части мозжечка. 4. Оптомоторный рефлекс проявляется в том, что при движении в поле зрения человека структурированного зрительного поля происходит непроизвольное движение глазных яблок в сторону движения стимулов. Когда движение глазных яблок в орбитах исчерпывает свой диапазон (по 45 угл. градуса от среднего положения), глаза скачкообразно возвращаются в исходное состояние. Совместная работа трех систем (вестибулоокулярной, оптомоторной и прослеживания) обеспечивает латентный период реакции глазного яблока, не превышающий 80 мс. Другими словами, человек способен следить за объектами, движущимися со скоростью около 300 угл. град, в секунду. 5. Шейные рефлексы на глаза. Если у животного (например, у кролика) удалить лабиринты, а затем поворачивать его голову, то можно получить все те перемещения глаз, которые описаны выше. Но в данном случае движения глаз осуществляются исключительно за счет шейных рефлексов. Положение глаз, вызванное этими рефлексами (при изменении положения головы или туловища), в конечном итоге приводит к тому, что изображение на сетчатках обоих глаз остается стабильным. В норме это достигается только совместным действием лабиринтных и шейных рефлексов. Шейноокулярный рефлекс довольно плохо выражен у интактных взрослых млекопитающих и практически отсутствует в норме у человека. 6. Шейный рефлекс на конечности. Если поднимать или опускать голову животного в атлантоокципитальном суставе, то вызываемые при этом рефлексы на передние и задние конечности действуют противоположным образом: при опускании головы активируются флексоры передних конечностей и экстензоры задних, при поднимании головы активируются экстензоры передних конечностей и флексоры задних (см. рис. 5.8). Сравнивая тонические рефлексы, можно видеть, что шейные и лабиринтные рефлексы на передние конечности действуют как синергисты, а на задние — как антагонисты. 7. Рефлекс перераспределения тонуса конечностей при поворотах головы. Если голову децеребрированной кошки (удалены большие полушария), лежащей на животе, поворачивать, то повышается тонус экстензорной мускулатуры в «челюстных» конечностях (конечности той половины туловища, к которым поворачивается нос), уменьшается и увеличивается — в «черепных» (конечности, к которым в этом случае поворачивается череп). Такое перераспределение тонуса для четвероногих животных оправдано соответствующим перенесением центра тяжести, и для того чтобы выдержать увеличенный вес в соответствующих конечностях, увеличивается их разгибательный (экстензорный) тонус. Можно получать все описанные шейные тонические рефлексы, если фиксировать позвоночный столб и перемещать только переднюю часть шеи. У здорового взрослого человека шейные и лабиринтные рефлексы не проявляются, но они отчетливо выступают в патологических состояниях (гидроцефалия, опухоли мозга, тяжелые мозговые травмы и пр.). Например, в этих условиях обнаруживаются тонические шейные рефлексы. Если поворачивать голову влево, чтобы нос приближался к левому плечу, то левые конечности становятся «челюстными», а правые — «черепными», поэтому повышается разгибательный тонус в обеих конечностях левой половины тела и уменьшается — справа. 8. Оптический установочный рефлекс играет особенно важную роль в правильной установке головы у высших млекопитающих. Например, обезьяна с разрушенным лабиринтом может правильно устанавливать голову, но если надеть ей колпак, закрывающий глаза, т.е. лишить зрения, она уже не сможет правильно устанавливать голову в пространстве. Таким образом, в организме животных взаимодействует ряд установочных рефлексов, благодаря которым обеспечивается нормальное положение головы и тела. Описанные установочные рефлексы осуществляются нервными центрами на уровне не выше среднего мозга. 9. Лифтная реакция представляет собой рефлекс на прямолинейное ускорение. Рефлекс состоит в том, что при поднятии площадки, на которой находится животное, тонус экстензоров снижается и конечности подгибаются; при опускании площадки развивается противоположная реакция: экстензоры активируются и конечности разгибаются. Эта реакция легко вызывается и у человека при подъеме или опускании в скоростном лифте.
Puc. 5.14. Свободное падение кошки из положения на спине (по Магнусу, 1924).
Описанные выше установочные рефлексы легко наблюдать на интактной кошке, которая свободно падает из положения на спине (рис. 5.14). Прежде всего активируются лабиринтные рефлексы на голову, благодаря которым она поворачивается по направлению к нормальному положению. К этой реакции присоединяется шейный установочный рефлекс, вследствие чего за головой следует тело, сначала грудной отдел и только затем таз. Происходит своего рода штопорообразное движение животного в пространстве, начинающееся с головы. При этом передние конечности разгибаются. Дальнейшее прямолинейное смещение головы в пространстве при свободном падении вызывает разгибание задних конечностей. Последнее можно объяснить лабиринтным рефлексом на падение (линейное ускорение). Благодаря совокупному действию этих рефлексов голова и тело кошки принимают нормальное положение, конечности тонически разгибаются и готовы принять вес тела, когда животное достигнет земли. Таким образом, положение тела и головы в пространстве и по отношению друг к другу принадлежит к наиболее застрахованным и автоматически выполняемым реакциям ЦНС. Благодаря взаимной деятельности большого количества разнородных рефлексов осуществляется единая конечная цель — правильная установка головы и тела в пространстве. Статокинетические и статические реакции взаимно дополняют друг друга: благодаря кинетическим реакциям выполняется движение, приводящее отдельные части тела в такое положение, в котором они потом удерживаются статическими рефлексами. Глазодвигательная система млекопитающих. В эволюции наземных позвоночных животных возникла необходимость сохранять неподвижность изображения на сетчатке не только в состоянии покоя или медленных движений, но также во время локомоции. Эволюция глазодвигательной системы у этих животных происходила в тесном взаимодействии с эволюцией зрительной системы. С точки зрения эволюции глазодвигательной системы всех позвоночных животных можно условно разделить на две группы: «фовеальные» животные, сетчатка которых имеет fovea и они обладают достаточно острым зрением, и «афовеальные» животные — с недостаточно острым зрением. У «фовеальных» животных (к ним относится и человек) глаза расположены во фронтальной плоскости, у них есть бинокулярное зрение. Зрительная ямка, или fovea, — это небольшая область в сетчатке с повышенной плотностью фоторецепторов, причем только в этой области находятся колбочки — фоторецепторы дневного (и цветового) зрения. Глазодвигательная система млекопитающих, которая сформировалась как необходимое дополнение зрительной системы, позволяет рассматривать зрительный мир. С точки зрения физиологии непрерывно решается задача — движением глаз поместить на зрительную ямку наиболее значимые части изображения. Что будет, если отключить fovea? Это происходит с каждым из нас при снижении уровня освещенности, например вечером. Известно, что мы в этих условиях не видим мелких предметов, не можем читать. Для такого зрения движения глаз не обязательны. Действительно, у животных с невыраженной fovea, например собаки, коровы, быстрые движения глаз отсутствуют. Глазодвигательная система млекопитающих выполняет следующие задачи: I) сохраняет неподвижным изображение внешнего мира на сетчатке во время движения животного относительно этого мира; 2) выделяет во внешнем мире некоторые объекты, помещает их в зоне сетчатки с высоким разрешением (например, fovea у человека) и прослеживает их движениями глаз и головы; 3) осуществляет скачкообразные (саккадические) перемещения взора для сканирования (рассматривания) внешнего мира. «Афовеальные» животные, например кошка, собака, кролик и др., не заботятся о положении изображения на сетчатке, и вестибулярная и оптокинетическая системы только предотвращают скольжение изображения по сетчатке. У человека в связи с изложенными выше требованиями, кроме вестибулоокулярной и оптокинетической систем, имеется также система прослеживания, вергенции и саккад. У него развились также рефлекторные механизмы содружественного движения глаз и головы. Саккадная система в полной мере развита только у обезьян и человека. Именно у них есть бинокулярное зрение и есть «фовеальное» зрение. Саккады — это быстрые содружественные отклонения глаз в быструю фазу вестибулярного и оптомоторного нистагмов, начальная фаза реакции прослеживания, когда скачком глаза «захватывается» движущаяся зрительная цель при зрительном обследовании внешнего мира. Структуры мозга, обеспечивающие быстрые скачкообразные движения глаз — саккады, находятся в ретикулярной формации мозгового ствола. Однако высший центр произвольного управления движениями глаз локализован в коре лобной доли мозга и называется фронтальным глазодвигательным полем (8-е и 10-е поля). Активность нейронов мозгового ствола. Схема, представленная на рис. 5.15, показывает связи и вероятные события, приводящие к саккаде глаза. Важными элементами этой схемы являются пачечные нейроны и нейроны-паузы. Во время фиксации взора клетки-паузы разряжаются тонически с высокой частотой; так как тормозные связи от них идут к тормозным и возбудительным пачечным нейронам, это должно предотвращать формирование пачки импульсов. Предполагают, что возбудительные пачечные нейроны проецируются к тоническим нейронам и происходит интегрирование их активности, в результате чего образуется сигнал, необходимый для поддержания глаза в эксцентричном положении (так называемый нейронный интегратор). В действительности нейронный интегратор, по-видимому, имеет большую протяженность. Установлено, что разрушение в области мозгового ствола и мозжечка
Рис. 5.15. Схема нейронного генератора саккад мозгового ствола. Слева — схема активности нейронов, справа — нейронная сеть, обеспечивающая саккаду.
приводит к параличу взора, т. е. неспособности поддерживать эксцентричное положение взора. По логике схемы стволового генератора запускающий сигнал формируется клетками-паузами. Установлено, что они получают афферентацию из средних и глубоких слоев верхнего двухолмия. Электрофизиологическими методами также было показано, что клетки-паузы могут возбуждаться моносинаптически на электрическую стимуляцию нижних слоев верхнего двухолмия. Таким образом, нейронная схема генератора саккады «работает» по принципу «спускового крючка» — клетка-пауза постоянно тормозит пачечные нейроны. Как только из верхнего двухолмия затормаживается разряд клетки-паузы, в этот интервал растормаживается пачечный нейрон и этот сигнал подается на соответствующие мотонейроны, что и приводит к саккаде. На основании электрофизиологических данных было высказано предположение, что пачечные нейроны со спонтанным разрядом являются промежуточным звеном между нижними слоями переднего двухолмия и парамедианной ретикулярной формацией моста. По мнению исследователей, эти нейроны могут получать конвергентные входы из ряда источников, в том числе из фронтального глазодвигательного поля. Известно, что дефицит саккад возникает только при одновременном разрушении переднего двухолмия и фронтального глазодвигательного поля. Вопросы 1. Уровни построения движений в нервной системе человека. 2. Спинной мозг — рефлекторный уровень построения движений. 3. Строение серого вещества спинного мозга. 4. Рефлексы мозгового ствола. 5. Нейрофизиология глазодвигательных реакций. Литература Бернштейн Н. Л. О построении движений. М.: Изд. АН СССР, 1970. Магнус Р. Установка тела. М.: Изд. АН СССР, 1962. Шульговский В. В. Физиология целенаправленного поведения млекопитающих. М.: Изд-во Моск. ун-та, 1993.
ФИЗИОЛОГИЯ МОЗЖЕЧКА Мозжечок, или малый мозг, представляет собой надсегментарную структуру, расположенную над продолговатым мозгом и мостом, позади больших полушарий мозга. Мозжечок состоит из нескольких частей, различных по происхождению в эволюции позвоночных животных. У человека мозжечок состоит из двух полушарий, находящихся по бокам от червя. К филогенетически более древней части мозжечка млекопитающих относят переднюю долю и флоккулонодулярную часть задней доли. Эти структуры мозжечка преимущественно связаны со спинным мозгом и вестибулярным аппаратом, тогда как полушария в основном получают информацию от мышечных и суставных рецепторов, а также от зрительного и слухового анализаторов. На рис. 5.16 представлена схема мозжечка млекопитающего
Рис. 5.16. Схема коры мозжечка (вид сверху). 1 — флоккулонодулярная доля; 2 — задняя доля; 3 — передняя доля; 4 — горизонтальная борозда.
(см. приложение 6), отражающая плотность вестибулярных, проприоцептивных (от мышц, сухожилий и суставов) и корковых афферентных проекций в различные зоны мозжечка. Согласно этой классификации кора мозжечка делится на три области: 1) архицеребеллум (старый мозжечок) — флоккулонодулярная доля (долька X); в ней оканчиваются преимущественно вестибулярные афференты и волокна от вестибулярных ядер; вестибулярные волокна проецируются также частично в язычок (lingula — долька I) и каудальную часть втулочки (uvula — долька IX), которые обычно относят также к архицеребеллуму; 2) палеоцеребеллум (древний мозжечок) включает переднюю долю (дольки II—V), простую дольку (долька VI) и заднюю часть корпуса мозжечка (дольки VIII—IX); палеоцеребеллум тесно связан со спинным мозгом, а также имеет двусторонние связи с сенсомоторной областью коры больших полушарий; 3) неоцеребеллум (новый мозжечок) включает среднюю часть корпуса мозжечка (долька VII и частично дольки VI и VIII), которая получает информацию от коры больших полушарий, а также
Рис. 5.17. Схема основных связей клеток Пуркинье коры мозжечка. Д — зубчатое ядро; Пр — промежуточное (округлое и пробковидное) ядро; Ш — ядро шатра; VL — вентролатеральное ядро таламуса; Кр.я. — красное ядро; В лат. — латеральное вестибулярное ядро (Дейтерса); РФ — ретикулярная формация мозгового ствола; Π — пирамидный нейрон двигательной коры; ВСТ — вестибулоспинальный тракт; РСТ — руброспинальный тракт; ПТ — пирамидный тракт; РетП — ретикулоспинальный путь.
от слуховых и зрительных рецепторов. Обратите внимание, что основная часть полушарий мозжечка принадлежит новому мозжечку, который лучше всего развит у человека. В толще мозжечка находятся три пары ядер: зубчатое, расположенное латерально; ядро шатра — медиально; пробковидное и округлое ядра — между ними. Единственным эфферентным выходом из коры мозжечка являются аксоны клеток Пуркинье, образующие синапсы с нейронами внутримозжечковых ядер и нейронами латерального вестибулярного ядра (рис. 5.17). Тесная связь ядра Дейтерса с корой мозжечка дает основание рассматривать его функционально как внутримозжечковое ядро. Все остальные образования головного и спинного мозга не получают прямых эфферентов из коры мозжечка. Ядра шатра посылают волокна к ядрам Дейтерса и к ретикулярной формации продолговатого мозга. Из области ретикулярной формации, где оканчиваются пути от мозжечка, берет начало ретикулоспинальный путь. Промежуточные ядра посылают аксоны в средний мозг, в том числе к красному ядру. Мощные пучки волокон, образованные преимущественно аксонами нейронов зубчатого ядра, направляются к вентролатеральному ядру таламуса, где происходит синаптическое переключение, и аксоны постсинаптических нейронов идут в моторную область коры больших полушарий; часть аксонов направляется к базальным ядрам. Таким образом, мозжечок не имеет самостоятельных двигательных систем, но образует обширные связи со всеми моторными системами: кортикоспинальной (пирамидной), руброспинальной, ретикулоспинальной, вестибулоспинальной, а также с полосатым телом. Афферентные и эфферентные волокна мозжечка собраны в три пары массивных волокнистых пучков, известных как мозжечковые ножки. Афферентные волокна входят в мозжечок в основном через нижние и средние ножки мозжечка. Эфферентные волокна проходят преимущественно через верхние ножки. Однако имеются исключения: некоторая часть спинно-мозжечковых путей входит через верхние ножки, а некоторые эфферентные волокна от флоккулонодулярной доли и ядра шатра проходят через нижние ножки. Ядра шатра направляют волокна через нижние ножки к вестибулярным ядрам и ретикулярной формации продолговатого мозга и моста. Промежуточные и зубчатые ядра посылают волокна через верхние ножки преимущественно к среднему мозгу и таламусу, особенно к красному ядру. Основная часть церебелло-таламических волокон отходит от зубчатых ядер. Эти волокна проецируются также к красному ядру, полосатому телу. Таким образом, влияния мозжечка на спинальные мотонейроны осуществляются через вестибулоспинальные и ретикулоспинальные пути, а на прецентральную область коры — через вентролатеральное ядро таламуса. Строение коры мозжечка. Кора мозжечка различных представителей позвоночных, включая человека, построена по единому плану и состоит из трех слоев (см. приложение 6). Поверхностный, или молекулярный, слой содержит разветвления дендритов клеток Пуркинье и параллельные волокна. Клетки Пуркинье имеют уплощенный дендрит, ориентированный параллельно сагиттальным зонам долек (folia) мозжечка. Дендриты и аксоны звездчатых клеток в молекулярном слое расположены таким же образом, тогда как параллельные волокна ориентированы строго трансверзально (перпендикулярно) по отношению к фоллиуму и сагиттальному направлению мшистых волокон. У кошки, например, параллельные волокна имеют среднюю длину 2 мм {диапазон до 5-7 мм). Одно параллельное волокно пересекает около 700 дендритов клеток Пуркинье. Проекции мшистых и лазающих волокон в коре мозжечка организованы в виде сагиттальных полосок. В нижней части молекулярного слоя находятся также тела корзинчатых клеток, аксоны которых идут перпендикулярно направлению листка коры мозжечка и оплетают тела и начальные сегменты аксонов клеток Пуркинье. В молекулярном слое имеется также небольшое число звездчатых клеток. Самый нижний, гранулярный слой содержит клетки-зерна, или гранулярные клетки. От тела клетки-зерна отходит 4-7 коротких дендритных отростков, с которыми мшистые волокна образуют синапсы. Аксон клетки-зерна поднимается вертикально вверх и в молекулярном слое Т-образно ветвится, образуя параллельные волокна. Плоскость его ветвления перпендикулярна плоскости ветвления дендритов клеток Пуркинье. В гранулярном слое находятся клетки Гольджи, аксоны которых восходят в молекулярный слой. Ганглиозный слой находится между описанными выше молекулярным и гранулярным слоями и содержит тела клеток Пуркинье. Афферентный вход в кору мозжечка осуществляется в основном через две системы волокон: лазающие и моховидные, или мшистые. Каждая клетка Пуркинье получает вход только от одного лазающего волокна (медиатор — аспартат), которое заканчивается на начальной (проксимальной) части дендрита. Лазающие волокна представляют собой аксоны нейронов, тела которых находятся в нижних оливах. По этому входу оказывается мощное возбуждающее действие: клетка Пуркинье деполяризуется на 10—15 мс, и в ней развиваются кальцийзависимые потенциалы действия. За потенциалом действия наступает следовая деполяризация, которая возникает вследствие активации кальцийзависимой калиевой проводимости соматической мембраны. После следовой деполяризации наступает следовая гиперполяризация. По этой причине клетка Пуркинье может возбуждаться по этому входу не более 1—2 раз в 1 с. |
||
|
|
Последнее изменение этой страницы: 2018-04-12; просмотров: 522. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |