|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
ГЛИЯ — МОРФОЛОГИЯ И ФУНКЦИЯГоловной мозг человека состоит из сотен миллиардов клеток, причем нервные клетки (нейроны) не составляют большинство. Большая часть объема нервной ткани (до 9/10 в некоторых областях мозга) занята клетками глии (от греч. склеивать). Дело в том, что нейрон выполняет в нашем организме гигантскую очень тонкую и трудную работу, для чего необходимо освободить такую клетку от
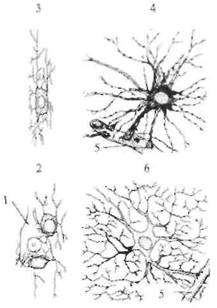
Рис. 2.2. Типы клеток глии головного мозга. 1 — нейрон; 2 — периневральный олигодендроцит; 3 — олигодендроцит в белом веществе; 4 — фиброзный астроцит; 5 — кровеносный сосуд; 6 — протоплазматический астроцит.
будничной деятельности, связанной с питанием, удалением шлаков, защитой от механических повреждений и т. д. — это обеспечивается другими, обслуживающими клетками, т.е. клетками глии (рис. 2.2). В головном мозге выделяют три типа клеток глии: микроглию, олигодендроглию и астроглию, каждая из которых обеспечивает только ей предназначенную функцию. Клетки микроглии участвуют в образовании мозговых оболочек, олигодендроглии — в образовании оболочек (миелиновых чехлов) вокруг отдельных отростков нервных клеток. Миелиновые оболочки вокруг периферических нервных волокон образуются специальными глиальными клетками — шванновскими клетками. Астроциты находятся вокруг нейронов, обеспечивая их механическую защиту, а кроме того, доставляют в нейрон питательные вещества и убирают шлаки. Клетки глии обеспечивают также электрическую изоляцию отдельных нейронов от воздействия других нейронов. Важной особенностью клеток глии является то, что в отличие от нейронов они сохраняют способность делиться на протяжении всей своей жизни. Это деление в некоторых случаях приводит к опухолевым заболеваниям головного мозга человека. Нервная клетка настолько специализирована, что утеряла способность к делению. Таким образом, нейроны нашего мозга, однажды образовавшись из клеток-предшественников (нейробластов), живут с нами всю нашу жизнь. На этом длительном пути мы только теряем нейроны нашего мозга.  НЕЙРОН Нейрон является главной клеткой центральной нервной системы. Формы нейронов чрезвычайно многообразны, но основные части неизменны у всех типов нейронов. Нейрон состоит из следующих частей: сомы (тела) и многочисленных разветвленных отростков. У каждого нейрона есть два типа отростков: аксон, по которому возбуждение передается от нейрона к другому нейрону, и многочисленные дендриты (от греч. дерево), на которых заканчиваются синапсами (от греч. контакт) аксоны от других нейронов. Нейрон проводит возбуждение только от дендрита к аксону. Основным свойством нейрона является способность возбуждаться (генерировать электрический импульс) и передавать (проводить) это возбуждение к другим нейронам, мышечным, железистым и другим клеткам. На рис. 2.3 показана схема нейрона, на которой легко прослеживаются его основные части. Нейроны разных отделов мозга выполняют очень разнообразную работу, и в соответствии с этим форма нейронов из разных частей головного мозга также многообразна (рис. 2.4). Нейроны, расположенные на выходе нейронной сети какой-то структуры, имеют длинный аксон, по которому возбуждение покидает данную мозговую структуру. Например, нейроны двигательной коры головного мозга, так называемые пирамиды Беца (названные в честь киевского анатома Б. Беца, впервые их описавшего в середине XIX века), имеют у человека аксон около 1 м, он соединяет двигательную кору больших полушарий с сегментами спинного мозга. По этому аксону передаются «двигательные команды», например «пошевелить пальцами ноги». Как возбуждается нейрон? Основная роль в этом процессе принадлежит мембране, которая отделяет цитоплазму клетки от окружающей среды. Мембрана нейрона, как и любой другой клетки,
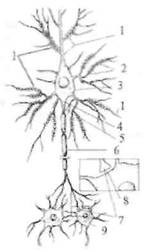
Рис. 2.3. Схема нейрона. 1 — дендрит; 2 — шипики; 3 — тело нейрона; 4 — дендрит; 5 — аксонный холмик; 6 — аксон; 7 — синаптическая щель; 8 — окончание аксона; 9 — постсинаптический нейрон.
Рис. 2.4. Нейроны различных структур головного мозга. 1 — клетка-зерно; 2 — двойная пирамида гиппокампа; 3 — пирамидная клетка; 4 — клетка Пуркинье; 5 — большая клетка ретикулярной формации: 6 — нейрон таламуса.
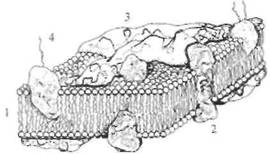
Рис. 2.5. Схема строения биологической мембраны. 1 — липиды; 2 — интегральные белки; 3 — периферические белки; 4 — гликопротеиды. Число полярных «головок» фосфолипидов приблизительно в 10 раз больше числа молекул интегральных белков.
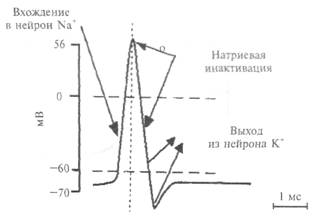
устроена очень сложно. В своей основе все известные биологические мембраны имеют однообразное строение (рис. 2.5): слой молекул белка, затем слой молекул липидов и еще один слой молекул белка. Вся эта конструкция напоминает два бутерброда, сложенных маслом друг к другу. Толщина такой мембраны составляет 7— 11 нм. Чтобы представить эти размеры, вообразите, что толщина вашего волоса уменьшилась в 10 тыс. раз. В такую мембрану встроены разнообразные частицы. Одни из них являются частицами белка и пронизывают мембрану насквозь (интегральные белки), они образуют места прохождения для ряда ионов; натрия, калия, кальция, хлора. Это так называемые ионные каналы. Другие частицы прикреплены на внешней поверхности мембраны и состоят не только из молекул белка, но и из полисахаридов. Это рецепторы для молекул биологически активных веществ, например медиаторов, гормонов и др. Часто в состав рецептора, кроме места для связывания специфической молекулы, входит и ионный канал. Главную роль в возбуждении нейрона играют ионные каналы мембраны. Эти каналы бывают двух видов: одни работают постоянно и откачивают из нейрона ионы натрия и накачивают в цитоплазму ионы калия. Благодаря работе этих каналов (их называют еще насосными каналами или ионным насосом), постоянно потребляющих энергию, в клетке создается разность концентраций ионов: внутри клетки концентрация ионов калия примерно в 30 раз превышает их концентрацию вне клетки, тогда как концентрация ионов натрия в клетке очень небольшая — примерно в 50 раз меньше, чем снаружи клетки. Свойство мембраны постоянно поддерживать разность ионных концентраций между цитоплазмой и окружающей средой характерно не только для нервной, но и для любой клетки организма. В результате между цитоплазмой и внешней средой на мембране клетки возникает потенциал: цитоплазма клетки заряжается отрицательно на величину около -70мВ относительно внешней среды клетки. Измерить этот потенциал можно в лаборатории стеклянным электродом, если в клетку ввести очень тонкую (меньше 1 мкм) стеклянную трубочку, заполненную раствором соли. Стекло в таком электроде играет роль хорошего изолятора, а раствор соли — проводника. Электрод соединяют с усилителем электрических сигналов и на экране осциллографа регистрируют этот потенциал. Оказывается, потенциал порядка —70 мВ сохраняется в отсутствие ионов натрия, но зависит от концентрации ионов калия. Другими словами, в создании этого потенциала участвуют только ионы калия, в связи с чем этот потенциал получил название «калиевый потенциал покоя», или просто «потенциал покоя·». Таким образом, это потенциал любой покоящейся клетки нашего организма, в том числе и нейрона. ВОЗБУЖДЕНИЕ НЕЙРОНА Нейрон в отличие от других клеток способен возбуждаться. Под возбуждением нейрона понимают генерацию нейроном потенциала действия. Основная роль в возбуждении принадлежит другому типу ионных каналов, при открытии которых ионы натрия устремляются в клетку. Напомним, что благодаря постоянной работе насосных каналов концентрация натриевых ионов вне клетки примерно в 50 раз больше, чем в клетке, поэтому при открытии натриевых каналов ионы натрия устремляются в клетку, а ионы калия через открытые калиевые каналы начинают выходить из клетки. Для каждого типа ионов — натрия и калия — имеется свой собственный тип ионного канала. Движение ионов по этим каналам происходит по концентрационным градиентам, т.е. из места высокой концентрации в место с более низкой концентрацией. Ответим на вопрос: как ионные каналы открываются и закрываются? В покоящемся нейроне натриевые каналы мембраны закрыты и на мембране, как это уже описывалось выше, регистрируется потенциал покоя порядка —70 мВ (отрицательность в цитоплазме). Если потенциал мембраны деполяризовать (уменьшить
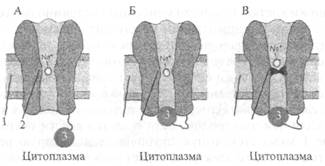
Рис. 2.6. Работа натриевого потенциалзависимого канала. А — канал открыт; Б — канал инактивирован; В — канал инактивирован и закрыт; 1 — мембрана; 2 — потенциалзависимый канал; 3 — инактивирующая частица.
поляризацию мембраны) примерно на 10 мВ, натриевый ионный канал открывается (рис. 2.6). Действительно, в канале имеется своеобразная заслонка, которая реагирует на потенциал мембраны, открывая этот канал при достижении потенциала определенной величины. Такой канал называется потенциалзависимым. Как только канал открывается, в цитоплазму нейрона устремляются из межклеточной среды ионы натрия, которых там примерно в 50 раз больше, чем в цитоплазме. Такое движение ионов является следствием простого физического закона: ионы движутся по концентрационному градиенту. Таким образом, в нейрон поступают ионы натрия, они заряжены положительно. Другими словами, через мембрану будет протекать входящий ток ионов натрия, который будет смещать потенциал мембраны в сторону деполяризации, т. е. уменьшать поляризацию мембраны. Чем больше ионов натрия войдет в цитоплазму нейрона, тем больше его мембрана деполяризуется. Потенциал на мембране будет увеличиваться, открывая все большее количество натриевых каналов. Но этот потенциал будет расти не бесконечно, а только до тех пор, пока не станет равным примерно +55 мВ. Этот потенциал соответствует присутствующим в нейроне и вне его концентрациям ионов натрия, поэтому его называют натриевым равновесным потенциалом. Вспомним, что в покое мембрана имела потенциал —70 мВ, тогда абсолютная амплитуда потенциала составит величину около 125 мВ. Мы говорим «около», «примерно» потому, что у клеток разного размера и типов этот
Рис. 2.7. Схема событий при возбуждении нейрона.
потенциал может несколько отличаться, что связано с формой этих клеток (например, количеством отростков), а также с особенностями их мембран. Все изложенное выше можно формально описать следующим образом. В покое клетка ведет себя как «калиевый электрод», а при возбуждении — как «натриевый электрод». Однако после того как потенциал на мембране достигнет своего максимального значения +55 мВ, натриевый ионный канал со стороны, обращенной в цитоплазму, закупоривается специальной белковой молекулой. Это так называемая «натриевая инактивация» (см. рис. 2.6); она наступает примерно через 0,5-1 мс и не зависит от потенциала на мембране. Мембрана становится непроницаемой для натриевых ионов. Для того чтобы потенциал мембраны вернулся к исходному состоянию — состоянию покоя, необходимо, чтобы из клетки выходил ток положительных частиц. Такими частицами в нейронах являются ионы калия. Они начинают выходить через открытые калиевые каналы. Вспомните, что в клетке в состоянии покоя накапливаются ионы калия, поэтому при открывании калиевых каналов эти ионы покидают нейрон, возвращая мембранный потенциал к исходному уровню (уровню покоя). В результате этих процессов мембрана нейрона возвращается к состоянию покоя (—70 мВ) и нейрон готовится к следующему акту возбуждения. Таким образом, выражением возбуждения нейрона является генерация на мембране нейрона потенциала действия. Его длительность
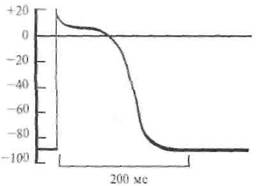
Рис. 2.8. Потенциал действия клетки желудочка сердца собаки.
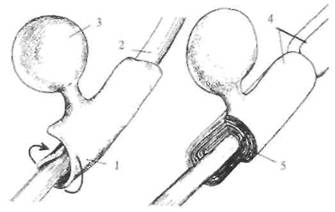
в нервных клетках составляет величину около 1/1000 с (1 мс). Описанная последовательность событий приведена на рис. 2.7. Подобные потенциалы действия могут возникать и в других клетках, назначение которых — возбуждаться и передавать это возбуждение другим клеткам. Например, сердечная мышца имеет в своем составе специальные мышечные волокна, обеспечивающие бесперебойную работу сердца в автоматическом режиме. В этих клетках также генерируются потенциалы действия (рис. 2.8). Однако они имеют затянутую, почти плоскую вершину, и длительность такого потенциала действия может затянуться до нескольких сот миллисекунд (сравните с 1 мс у нейрона). Такой характер потенциала действия мышечной клетки сердца физиологически оправдан, так как возбуждение сердечной мышцы должно быть длительным, чтобы кровь успела покинуть желудочек. С чем же связан такой затянутый потенциал действия у этого типа клетки? Оказалось, в мембране этих клеток натриевые ионные каналы не так быстро закрываются, как в нейронах, т. е. натриевая инактивация затянута. Как ясно из этого описания, возбуждение (потенциал действия) нейрона сменяется так называемым «покоем». Однако никакого покоя в этот период нет. Как уже указывалось выше, в мембране есть еще и насосные каналы, количество которых примерно в 10 раз больше ионных, и они постоянно работают, откачивая из цитоплазмы излишек ионов натрия и закачивая туда недостающие ионы калия. Благодаря неустанной работе этих каналов нейрон всегда готов к возбуждению. Описанный выше механизм возбуждения клетки (конечно, далеко не все клетки нашего организма способны возбуждаться) в основных чертах одинаков не только в нейронах и мышечных клетках человека, но и в аналогичных клетках других организмов. Например, в нейронах моллюсков, червей, крыс и обезьян при возбуждении происходят описанные выше последовательности событий. Более того, конструкция мембран, включая каналы, также примерно одинакова у всех организмов Земли. Как уже указывалось, каналы представляют собой белковые молекулы, «прошивающие» мембрану (одна часть молекулы находится в цитоплазме, а другая — во внеклеточной среде). Интересно, что эти белковые молекулы, образующие ионный или насосный каналы, не вечны, а постоянно заменяются на новые (примерно каждые несколько часов). Все это свидетельствует об очень большой динамичности структуры нейрона. Нейрон способен к возбуждению, которое состоит в том, что мембрана нейрона в состоянии покоя имеет потенциал порядка —70мВ (отрицательность β цитоплазме), а β состоянии возбуждения приобретает потенциал +55 мВ. Таким образом, абсолютная величина потенциала действия — около 125 мВ. Длительность потенциала действия нейрона составляет всего около 1 мс (1/1000 с). Далее это возбуждение (потенциал действия) должно передаться другому нейрону или какой-то другой клетке, например мышечной, железистой и др. ПРОВЕДЕНИЕ ВОЗБУЖДЕНИЯ Возбуждение в виде потенциала действия покидает тело нейрона по его отростку, который называется аксоном. Аксоны отдельных нейронов обычно объединяются в пучки — нервы, а сами аксоны в этих пучках называются нервными волокнами. Природа позаботилась, чтобы волокна максимально хорошо справлялись с функцией проведения возбуждения в виде потенциалов действия. Для этой цели отдельные нервные волокна (аксоны отдельных нейронов) имеют специальные чехлы, выполненные из хорошего электрического изолятора (см. рис. 2.3). Чехол прерывается примерно через каждые 0,5-1,5 мм; это связано с тем, что отдельные участки чехла образуются в результате того, что специальные клетки в очень ранний период развития организма (в основном еще до рождения) обволакивают небольшие участки аксона. На рис. 2.9 показано, как это происходит. В периферических нервах миелин
Рис. 2.9. Образование миелинового чехла на аксоне. 1 — наматывание слоев миелина; 2 — аксон: 3 — олигодендроцит; 4 — перехват Ранвье; 5 — слой миелиновой оболочки.
образуется клетками, которые получили название шванновских, а в головном мозге это происходит за счет клеток олигодендроглии. Этот процесс называется миелинизацией, так как в результате образуется чехол из вещества миелина, примерно на 2/3 состоящего из жира и являющегося хорошим электрическим изолятором. Исследователи придают очень большое значение процессу миелинизации в развитии мозга. Известно, что у новорожденного ребенка миелинизировано примерно 2/3 волокон головного мозга. Примерно к 12 годам завершается следующий этап миелинизации. Это соответствует тому, что у ребенка уже формируется функция внимания, он достаточно хорошо владеет собой. Вместе с тем полностью процесс миелинизации заканчивается только при завершении полового созревания. Таким образом, процесс миелинизации является показателем созревания ряда психических функций. В то же время известны заболевания нервной системы человека, которые связаны с демиелинизацией нервных волокон, что сопровождается тяжелыми страданиями. К самым известным относится рассеянный склероз. Это заболевание развивается незаметно и очень медленно, последствием является паралич движения. Почему же так важна миелинизация нервных волокон? Оказывается, миелинизированные волокна в сотни раз быстрее проводят
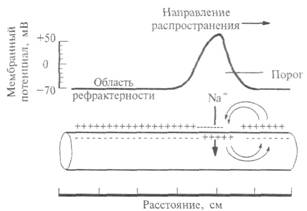
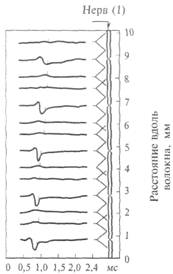
Рис. 2.10. Распространение возбуждения по немиелинизированному нервному волокну: после прохождения потенциала действия в нервном волокне возникает зона невозбудимости, или рефрактерности.
возбуждение, чем немиелинизированные, т. е. нейронные сети нашего мозга могут работать с большей скоростью, а значит, более эффективно. Поэтому не миелинизируются в нашем организме только самые тонкие волокна (менее 1 мкм в диаметре), которые проводят возбуждение к медленно работающим органам — кишечнику, мочевому пузырю и др. Как правило, не миелинизируются волокна, проводящие информацию о боли и температуре. Как происходит распространение возбуждения по нервному волокну? Вначале разберем случай немиелинизированного нервного волокна. На рис. 2.10 показана схема нервного волокна. Возбужденный участок аксона характеризуется тем, что мембрана, обращенная к аксоплазме, заряжается положительно относительно экстраклеточной среды. Невозбужденные (покоящиеся) участки мембраны волокна отрицательны внутри. Между возбужденным и невозбужденным участками мембраны возникает разность потенциалов и начинает протекать ток. На рисунке это отражено линиями тока, пересекающими мембрану со стороны аксоплазмы, — выходящий ток, который деполяризует соседний невозбуждениый участок волокна. Возбуждение движется по волокну только в одном направлении (показано стрелкой) и не может пойти в другую сторону, так как после возбуждения участка волокна в нем наступает рефрактерность — зона не возбудимости. Нам уже известно,
Рис. 2.11. Распространение возбуждения по миелинизированному нервному волокну: потенциалы действия возникают только в перехватах Ранвье.
что деполяризация приводит к открыванию потенциалзависимых натриевых каналов и в соседнем участке мембраны развивается потенциал действия. Затем натриевый канал инактивируется и закрывается, что и приводит к зоне невозбудимости волокна. Эта последовательность событий повторяется для каждого соседнего участка волокна. На каждое такое возбуждение тратится определенное время. Специальные исследования показали, что скорость проведения возбуждения немиелинизированных волокон пропорциональна их диаметру: чем больше диаметр, тем выше скорость движения импульсов. Например, немиелинизированные волокна, проводящие возбуждение со скоростью 100—120 м/с, должны иметь диаметр около 1000 мкм (1 мм). У млекопитающих животных природа сохранила немиелинизированными только те волокна, которые проводят возбуждение о боли, температуре, управляют медленно работающими внутренними органами — мочевым пузырем, кишечником и пр. Практически все нервные волокна в центральной нервной системе человека имеют миелиновые чехлы. На рис. 2.11 показано, что если вдоль волокна, покрытого миелином, регистрировать прохождение возбуждения, то потенциал действия возникает только в перехватах Ранвье. Оказывается, миелин, являясь хорошим электрическим изолятором, не пропускает выхода линий тока от предшествующего возбужденного участка. Выход тока в этом случае возможен только через те участки мембраны, которые находятся на стыке между двумя участками миелина. Напомним, что каждый участок образован только одной клеткой, поэтому это стыки между двумя клетками, образующими соседние участки миелиновой оболочки. Мембрана аксона между двумя соседними миелиновыми чехлами оказывается не покрытой миелином (так называемый перехват Ранвье). Благодаря такому устройству мембрана волокна возбуждается только в местах перехватов Ранвье. Вследствие этого потенциал действия (возбуждение) как бы перескакивает через участки изолированной мембраны. Другими словами, возбуждение движется скачками от перехвата к перехвату. Это похоже на те волшебные сапоги-скороходы, которые надевал кот в известной сказке, мгновенно переносясь из одного места в другое. СИНАПС Как передается возбуждение — от одного нейрона другому или от нейрона, например, на мышечное волокно? Этой проблемой интересуются не только профессиональные нейробиологи, но и врачи, особенно фармакологи. Знание биологических механизмов необходимо для лечения некоторых заболеваний, а также для создания новых лекарств и препаратов. Дело в том, что одними из основных мест воздействия этих веществ на организм человека являются места передачи возбуждения с одного нейрона на другой (или на другую клетку, например клетку сердечной мышцы, стенки сосудов и пр.). Отросток нейрона аксон направляется к другому нейрону и образует на нем контакт, который называют синапсом (в переводе с греческого — контакт; см. рис. 2.3). Именно синапс хранит многие тайны мозга. Нарушение этого контакта, например, веществами, блокирующими его работу, приводит к тяжелейшим последствиям для человека. Это место приложения действия наркотиков. Примеры будут приведены ниже, а сейчас рассмотрим, как устроен и как работает синапс. Трудности этого исследования определяются тем, что сам синапс очень маленький (его диаметр не более 1 мкм). Один нейрон получает такие контакты, как правило, от нескольких тысяч (3— 10 тыс.) других нейронов. Каждый синапс надежно закрыт специальными
Рис. 2.12. Синапс (схема). 1 — везикулы; 2 — митохондрии; 3 — пресинаптическая мембрана; 4 — синаптическая щель; 5 — ионный канал; 6 — постсинаптическая мембрана.
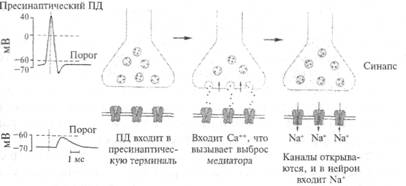
клетками глии, поэтому исследовать его очень непросто. На рис. 2.12 показана схема синапса, как это представляет себе современная наука. Несмотря на свою миниатюрность, он устроен весьма сложно. Одним из его основных компонентов являются пузырьки, которые находятся внутри синапса. Эти пузырьки содержат биологически очень активное вещество, которое называется нейротрансмиттером, или медиатором (передатчиком). Вспомним, что нервный импульс (возбуждение) с огромной скоростью продвигается по волокну и подходит к синапсу. Этот потенциал действия вызывает деполяризацию мембраны синапса (рис. 2.13), однако это не приводит к генерации нового возбуждения (потенциала действия), а вызывает открывание специальных ионных каналов, с которыми мы еще не знакомы. Эти каналы пропускают ионы кальция внутрь синапса. Ионы кальция играют очень большую роль в деятельности организма. Специальная железа внутренней секреции — паращитовидная (она находится поверх щитовидной железы) — регулирует содержание кальция в организме. Многие заболевания связаны с нарушением обмена кальция в организме. Например, его недостаток приводит к рахиту у маленьких детей.
Рис. 2.13. Последовательность событий при возбуждении синапса.
Каким образом кальций участвует в работе синапса? Попадая в цитоплазму синаптического окончания, кальций входит в связь с белками, образующими оболочку пузырьков, в которых хранится медиатор. В конечном итоге мембраны синаптических пузырьков сжимаются, выталкивая свое содержимое в синаптическую щель. Этот процесс очень напоминает сокращение мышечного волокна в мышце, во всяком случае, эти два процесса имеют одинаковый механизм на молекулярном уровне. Таким образом, связывание кальция белками оболочки пузырька приводит к ее сокращению и содержание пузырька впрыскивается (экзоцитоз) в щель, которая отделяет мембрану одного нейрона от мембраны другого. Эта щель называется синоптической щелью. Из описания должно быть ясно, что возбуждение (электрический потенциал действия) нейрона в синапсе превращается из электрического импульса в импульс химический. Другими словами, каждое возбуждение нейрона сопровождается выбросом в окончании его аксона порции биологически активного вещества — медиатора. Далее молекулы медиатора связываются с специальными белковыми молекулами, которые находятся на мембране другого нейрона. Эти молекулы называются рецепторами. Рецепторы устроены уникально и связывают только один тип молекул. В некоторых описаниях указывается, что они подходят, как «ключ к замку» (ключ подходит только к своему замку). Рецептор состоит из двух частей. Одну можно назвать «узнающим центром», другую — «ионным каналом». Если молекулы медиатора заняли определенные места (узнающий центр) на молекуле рецептора, то ионный канал открывается и ионы начинают входить в клетку (ионы натрия) или выходить (ионы калия) из клетки. Другими словами, через мембрану протекает ионный ток, который вызывает изменение потенциала на мембране. Этот потенциал получил название постсинаптического потенциала (рис. 2.13). Очень важным свойством описанных ионных каналов является то, что количество открытых каналов определяется количеством связанных молекул медиатора, а не потенциалом на мембране, как в случае с электровозбудимой мембраной нервного волокна. Таким образом, постсинаптические потенциалы имеют свойство градуальности: амплитуда потенциала определяется количеством молекул медиатора, связанного рецепторами. Благодаря этой зависимости амплитуда потенциала на мембране нейрона развивается пропорционально количеству открытых каналов. На мембране одного нейрона могут одновременно находиться два вида синапсов: тормозные и возбудительные. Все определяется устройством ионного канала мембраны. Мембрана возбудительных синапсов пропускает как ионы натрия, так и ионы калия. В этом случае мембрана нейрона деполяризуется. Мембрана тормозных синапсов пропускает только ионы хлора и гиперполяризуется. Очевидно, что если нейрон заторможен, потенциал мембраны увеличивается (гиперполяризация). Таким образом, нейрон благодаря воздействию через соответствующие синапсы может возбудиться или прекратить возбуждение, затормозиться. Все эти события происходят на соме и многочисленных отростках дендрита нейрона, на последних находится до нескольких тысяч тормозных и возбудительных синапсов. В качестве примера разберем, как действует в синапсе медиатор, который называется ацетилхолином. Этот медиатор широко распространен в головном мозге и в периферических окончаниях нервных волокон. Например, двигательные импульсы, которые по соответствующим нервам приводят к сокращению мышц нашего тела, оперируют ацетилхолином. Ацетилхолин был открыт в 30-х годах австрийским ученым О. Леви. Эксперимент был очень прост: изолировали сердце лягушки с подходящим к нему блуждающим нервом. Было известно, что электрическая стимуляция блуждающего нерва приводит к замедлению сокращений сердца вплоть до полной его остановки. О. Леви простимулировал блуждающий нерв, получил эффект остановки сердца и взял из сердца немного крови. Оказалось, что если эту кровь добавить в желудочек работающего сердца, то оно замедляет свои сокращения. Был сделан вывод: при стимуляции блуждающего нерва выделяется вещество, останавливающее сердце. Это и был ацетилхолин. Позже был открыт фермент, который расщеплял ацетилхолин на холин (жир) и уксусную кислоту, в результате чего прекращалось действие медиатора. Этим исследованием впервые была установлена точная химическая формула медиатора и последовательность событий в типичном химическом синапсе. Эта последовательность событий сводится к следующему. Потенциал действия, пришедший по пресинаптическому волокну к синапсу, вызывает деполяризацию, которая включает кальциевый насос, и ионы кальция поступают в синапс; ионы кальция связываются белками мембраны синаптических пузырьков, что приводит к активному опорожнению (экзоцитозу) пузырьков в синаптическую щель. Молекулы медиатора связываются (узнающим центром) соответствующими рецепторами постсинаптической мембраны, при этом открывается ионный канал. Через мембрану начинает протекать ионный ток, что приводит к возникновению на ней постсинаптического потенциала. В зависимости от характера открытых ионных каналов возникает возбудительный (открываются каналы для ионов натрия и калия) или тормозной (открываются каналы для ионов хлора) постсинаптический потенциал. Ацетилхолин весьма широко распространен в живой природе. Например, он находится в стрекательных капсулах крапивы, в стрекательных клетках кишечнополостных животных (например, пресноводной гидры, медузы) и пр. В нашем организме ацетилхолин выбрасывается в окончаниях двигательных нервов, управляющих мышцами, из окончаний блуждающего нерва, который управляет деятельностью сердца и других внутренних органов. Человек давно знаком с антагонистом ацетилхолина — это яд кураре, которым пользовались индейцы Южной Америки при охоте на животных. Оказалось, что кураре, попадая в кровь, вызывает обездвиживание животного, и оно погибает фактически от удушья, но кураре не останавливает сердце. Исследования показали, что в организме существуют два типа рецепторов к ацетилхолину: один успешно связывает никотиновую кислоту, а другой — мускарин (вещество, которое выделено из гриба рода Muscaris). На мышцах нашего тела находятся рецепторы никотинового типа к ацетилхолину, тогда как на сердечной мышце и нейронах головного мозга — рецепторы к ацетилхолину мускаринового типа. В настоящее время в медицине широко применяют синтетические аналоги кураре для обездвиживания больных во время сложных операций на внутренних органах. Применение этих средств приводит к полному параличу двигательной мускулатуры (связывается рецепторами никотинового типа), но не влияет на работу внутренних органов, в том числе сердца (рецепторы мускаринового типа). Нейроны головного мозга, возбуждаемые через мускариновые ацетилхолиновые рецепторы, играют большую роль в проявлении некоторых психических функций. Сейчас известно, что гибель таких нейронов приводит к старческому слабоумию (болезнь Альцгеймера). Другим примером, который должен показать важность именно рецепторов никотинового типа на мышце к ацетилхолину, может служить заболевание, называемое miastenia grevis (мышечная слабость). Это генетически наследуемая болезнь, т. е. ее происхождение связано с «поломками» генетического аппарата, которые передаются по наследству. Заболевание проявляется в возрасте ближе к половозрелости и начинается с мышечной слабости, которая постепенно усиливается и захватывает все более обширные группы мышц. Причиной этого недуга оказалось то, что организм больного вырабатывает белковые молекулы, которые прекрасно связываются ацетилхолиновыми рецепторами никотинового типа. Занимая эти рецепторы, они препятствуют связыванию с ними молекул ацетилхолина, выбрасываемых из синаптических окончаний двигательных нервов. Это и приводит к блокированию синаптического проведения к мышцам и, следовательно, к их параличу. Описанный на примере ацетилхолина тип синаптической передачи — не единственный в ЦНС. Второй тип синаптической передачи также широко распространен, например в синапсах, в которых медиаторами являются биогенные амины (дофамин, серотонин, адреналин и др.). В этом типе синапсов имеет место следующая последовательность событий. После того как образовался комплекс «молекула медиатора — рецепторный белок», активируется специальный мембранный белок (G-белок). Одна молекула медиатора при связывании с рецептором может активировать много молекул G-белка, и это усиливает эффект медиатора. Каждая активированная молекула G-белка в одних нейронах может открывать ионный канал, а в других — активировать внутри клетки синтез специальных молекул, так называемых вторичных посредников. Вторичные посредники могут запускать в клетке многие биохимические реакции, связанные с синтезом, например, белка, в этом случае возникновения электрического потенциала на мембране нейрона не происходит. Существуют и другие медиаторы. В головном мозге в качестве медиаторов «работает» целая группа веществ, которые объединены под названием биогенные амины. В середине прошлого столетия английский врач Паркинсон описал болезнь, которая проявлялась как дрожательный паралич. Это тяжелое страдание вызвано разрушением в мозге больного нейронов, которые в своих синапсах (окончаниях) выделяют дофамин — вещество из группы биогенных аминов. Тела этих нейронов находятся в среднем мозге, образуя там скопление, которое называется черной субстанцией. Исследования последних лет показали, что дофамин в мозге млекопитающих также имеет несколько типов рецепторов (в настоящеее время известно шесть типов). Другое вещество из группы биогенных аминов — серотонин (другое название 5-окситриптамин) — вначале было известно как средство, приводящее к подъему кровяного давления (сосудосуживающее). Обратите внимание, что это отражено в его названии. Однако оказалось, что истощение в головном мозге серотонина приводит к хронической бессоннице. В опытах на животных было установлено, что разрушение в мозговом стволе (задних отделах мозга) специальных ядер, которые известны в анатомии как ядра шва, приводит к хронической бессоннице и в дальнейшем гибели этих животных. Биохимическое исследование установило, что нейроны ядер шва содержат серотонин. У пациентов, страдающих хронической бессонницей, также обнаружено снижение концентрации серотонина в мозге. К биогенным аминам относят также адреналин и норадреналин, которые содержатся в синапсах нейронов автономной вегетативной нервной системы. Во время стресса под влиянием специального гормона — адренокортикотропного (подробнее см. ниже) — из клеток коры надпочечников в кровь также выбрасываются адреналин и норадреналин. МЕДИАТОРЫ НЕРВНОЙ СИСТЕМЫ Из вышеизложенного понятно, какое значение в функциях нервной системы играют медиаторы. В ответ на приход нервного импульса к синапсу происходит выброс медиатора; молекулы медиатора соединяются (комплементарно — как «ключ к замку») с рецепторами постсинаптической мембраны, что приводит к открыванию ионного канала или к активированию внутриклеточных реакций. Примеры синаптической передачи, рассмотренные выше, полностью соответствуют этой схеме. Вместе с тем благодаря исследованиям последних десятилетий эта довольно простая схема химической синаптической передачи значительно усложнилась. Появление иммунохимических методов позволило показать, что в одном синапсе могут сосуществовать несколько групп медиаторов, а не один, как это предполагали раньше. Например, в одном синаптическом окончании одновременно могут находиться синаптические пузырьки, содержащие ацетилхолин и норадреналин, которые довольно легко идентифицируются на электронных фотографиях (ацетилхолин содержится в прозрачных пузырьках диаметром около 50 нм, а норадреналин — в электронно-плотных диаметром до 200 нм). Кроме классических медиаторов, в синаптическом окончании могут находиться один или несколько нейропептидов. Количество веществ, содержащихся в синапсе, может доходить до 5—6 (своеобразный коктейль). Более того, медиаторная специфичность синапса может меняться в онтогенезе. Например, нейроны симпатических ганглиев, иннервирующие потовые железы у млекопитающих, исходно норадренергичны, но у взрослых животных становятся холинергичными. В настоящее время при классификации медиаторных веществ принято выделять: первичные медиаторы, сопутствующие медиаторы, медиаторы-модуляторы и аллостерические медиаторы. Первичными медиаторами считают те, которые действуют непосредственно на рецепторы постсинаптической мембраны. Сопутствующие медиаторы и медиаторы-модуляторы могут запускать каскад ферментативных реакций, которые, например, фосфорилируют рецептор для первичного медиатора. Аллостерические медиаторы могут участвовать в кооперативных процессах взаимодействия с рецепторами первичного медиатора. Долгое время за образец принимали синаптическую передачу по анатомическому адресу (принцип «точка — в точку»). Открытия последних десятилетий, особенно медиаторной функции нейропептидов, показали, что в нервной системе возможен принцип передачи и по химическому адресу. Другими словами, медиатор, выделяющийся из данного окончания, может действовать не только на «свою» постсинаптическую мембрану, но и за пределами данного синапса — на мембраны других нейронов, имеющих соответствующие рецепторы. Таким образом, физиологическая реакция
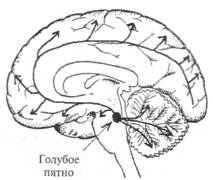
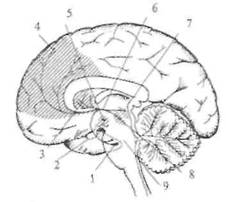
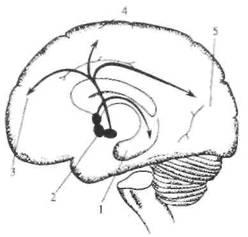
Рис. 2.14. Схема норадренергических путей в головном мозге. Основным источником норадренергических аксонов являются нейроны голубого пятна — nucl. cereleus (указано стрелкой) и прилежащих участков среднего мозга. Аксоны этих нейронов широко распространяются в стволе мозга, мозжечке, конечном мозге, а также в спинном мозге (на схеме не показано).
обеспечивается не точным анатомическим контактом, а наличием соответствующего рецептора на клетке-мишени. Собственно этот принцип был давно известен в эндокринологии, а исследования последних лет нашли ему более широкое применение. Все известные типы хеморецепторов на постсинаптической мембране разделяют на две группы. В одну группу входят рецепторы, в состав которых включен ионный канал, открывающийся при связывании молекул медиатора с «узнающим» центром. Рецепторы второй группы (метаботропные рецепторы) открывают ионный канал опосредованно (через цепочку биохимических реакций), в частности, посредством активации специальных внутриклеточных белков. Одними из самых распространенных являются медиаторы, принадлежащие к группе биогенных аминов. Эта группа медиаторов достаточно надежно идентифицируется микрогистологическими методами. Известны две группы биогенных аминов: катехоламины (дофамин, норадреналин и адреналин) и индоламин (серотонин). Функции биогенных аминов в организме весьма многообразны: медиаторная, гормональная, регуляция эмбриогенеза. Основным источником норадренергических аксонов являются нейроны голубого пятна и прилежащих участков среднего мозга (рис. 2.14). Аксоны этих нейронов широко распространяются в мозговом
Рис. 2.15. Дофаминергические системы головного мозга. 1 — энторинальная кора; 2 — гипофиз; 3 — хвостатое ядро; 4 — фронтальная кора; 5 — гипоталамус; 6 — медиальный пучок переднего мозга; 7 — черная субстанция; 8 — средний мозг (покрышка); 9 — мозговой ствол.
стволе, мозжечке, в больших полушариях. В продолговатом мозге крупное скопление норадренергических нейронов находится в вентролатеральном ядре ретикулярной формации. В промежуточном мозге (гипоталамусе) норадренергические нейроны наряду с дофаминергическими нейронами входят в состав гипоталамо-гипофизарной системы. Норадренергические нейроны в большом количестве содержатся в периферической нервной системе. Их тела лежат в симпатической цепочке и в некоторых интрамуральных ганглиях. Дофаминергические нейроны у млекопитающих находятся преимущественно в среднем мозге (так называемая нигро-неостриарная система), а также в гипоталамической области. Дофаминовые цепи мозга млекопитающих хорошо изучены. Известны три главные цепи, все они состоят из однонейронной цепочки. Тела нейронов находятся в мозговом стволе и отсылают аксоны в другие области головного мозга (рис. 2.15). Одна цепь очень проста. Тело нейрона находится в области гипоталамуса и отсылает короткий аксон в гипофиз. Этот путь входит в состав гипоталамо-гипофизарной системы и контролирует систему эндокринных желез. Вторая дофаминовая система также хорошо изучена. Это черная субстанция, многие клетки которой содержат дофамин. Аксоны этих нейронов проецируются в полосатые тела. Эта система содержит примерно 3/4 дофамина головного мозга. Она имеет решающее значение в регулировании тонических движений, Дефицит дофамина в этой системе приводит к болезни Паркинсона. Известно, что при этом заболевании происходит гибель нейронов черной субстанции. Введение L-DOPA (предшественника дофамина) облегчает у больных некоторые симптомы заболевания. Третья дофаминергическая система участвует в проявлении шизофрении и некоторых других психических заболеваний. Функции этой системы пока изучены недостаточно, хотя сами пути хорошо известны. Тела нейронов лежат в среднем мозге рядом с черной субстанцией. Они проецируют аксоны в вышележащие структуры мозга, мозговую кору и лимбическую систему, особенно к фронтальной коре, к септальной области и энторинальной коре. Энторинальная кора, в свою очередь, является главным источником проекций к гиппокампу. Согласно дофаминовой гипотезе шизофрении, третья дофаминергическая система при этом заболевании сверхактивна. Эти представления возникли после открытия веществ, снимающих некоторые симптомы заболевания. Например, хлорпромазин и галоперидол имеют разную химическую природу, но они одинаково подавляют активность дофаминергической системы мозга и проявление некоторые симптомов шизофрении, У больных шизофренией, в течение года получавших эти препараты, появляются двигательные нарушения, получившие название tardive dyskinesia (повторяющиеся причудливые движения лицевой мускулатуры, включая мускулатуру рта, которые больной не может контролировать). Серотонин почти одновременно открыли в качестве сывороточного сосудосуживающего фактора (1948) и энтерамина, секретируемого энтерохромаффиновыми клетками слизистой оболочки кишечника. В 1951 г. было расшифровано химическое строение серотонина и он получил новое название — 5-гидрокситриптамин. В организме млекопитающих он образуется гидроксилированием аминокислоты триптофана с последующим декарбоксилированием. 90% серотонина образуется в организме энтерохромаффиновыми клетками слизистой оболочки всего пищеварительного тракта. Внутриклеточный серотонин инактивируется моноаминоксидазой, содержащейся в митохондриях. Серотонин внеклеточного пространства окисляется перулоплазмином. Большая часть вырабатываемого серотонина связывается с кровяными пластинками и по кровяному руслу разносится по организму. Другая часть действует в качестве
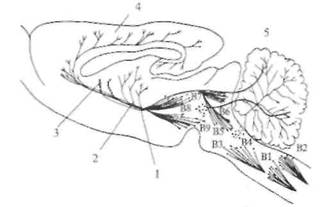
Рис. 2.16. Серотонинергичсские пути в головном мозге крысы (схема). В1-В9 — группы серотонинергических нейронов в области шва, аксоны которых проецируются в кору больших полушарий, гипоталамус и другие структуры; 1 — медиальный пучок переднего мозга; 2 — гипоталамус; 3 — хвостатое ядро; 4 — кора; 5 — мозжечок.
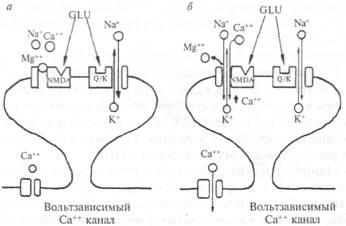
местного гормона, способствуя авторегулированию кишечной перистальтики, а также модулируя эпителиальную секрецию и всасывание в кишечном тракте. Серотонинергические нейроны широко распространены в центральной нервной системе (рис. 2.16). Они обнаруживаются в составе дорсального и медиального ядер шва продолговатого мозга, а также в среднем мозге и варолиевом мосту. Серотонинергические нейроны иннервируют обширные области мозга, включающие кору больших полушарий, гиппокамп, бледный шар, миндалину, область гипоталамуса. Интерес к серотонину был привлечен в связи с проблемой сна. При разрушении ядер шва животные страдали бессонницей. Сходный эффект оказывали вещества, истощающие хранилище серотонина в мозге. Самая высокая концентрация серотонина обнаружена в эпифизе (pineal gland). Серотонин в эпифизе превращается в мелатонин, который участвует в пигментации кожи, а также влияет у многих животных на активность женских гонад. Содержание как серотонина, так и мелатонина в эпифизе контролируется циклом свет—темнота через симпатическую нервную систему. Другую группу медиаторов ЦНС составляют аминокислоты. Уже давно известно, что нервная ткань с ее высоким уровнем метаболизма содержит значительные концентрации целого набора аминокислот (перечислены в порядке убывания): глутаминовой кислоты, глутамина, аспарагиновой кислоты, гамма-аминомасляной кислоты (ГАМК). Глутамат в нервной ткани образуется преимущественно из глюкозы. У млекопитающих больше всего глутамата содержится в конечном мозге и мозжечке, где его концентрация примерно в 2 раза выше, чем в стволе мозга и спинном мозге. В спинном мозге глутамат распределен неравномерно: в задних рогах он находится в большей концентрации, чем в передних. Глутамат является одним из самых распространенных медиаторов в ЦНС. Посте инаптические рецепторы к глутамату классифицируются в соответствии с аффинностью (сродством) к трем экзогенным агонистам — квисгулату, каинату и N-метил-D-аспартату (NMDA). Ионные каналы, активируемые квисгулатом и каина-том, подобны каналам, которые управляются никотиновыми рецепторами — они пропускают смесь катионов (Na+ и К+). Стимуляция NMDA-рецепторов имеет сложный характер активации: ионный ток, который переносится не только Na+ и К+, но также Са++ при открывании ионного канала рецептора, зависит от потенциала мембраны. Вольтзависимая природа этого канала определяется разной степенью его блокирования ионами Mg++ с учетом уровня мембранного потенциала. При потенциале покоя порядка —75 мВ ионы Mg++, которые преимущественно находятся в межклеточной среде, конкурируют с ионами Са++ и Na+ за соответствующие каналы мембраны (рис. 2.17). Вследствие того что ион Mg++ не может пройти через пору, канал блокируется всякий раз, как попадает туда ион Mg++. Это приводит к уменьшению времени открытого канала и проводимости мембраны. Если мембрану нейрона деполяризовать, то количество ионов Mg++, которые закрывают ионный канал, снижается и через канал беспрепятственно могут проходить ионы Са++, Na+ и К+. При редких стимуляциях (потенциал покоя изменяется мало) глутаматергического рецептора ВПСП возникает преимущественно за счет активации квисгулатных и каинатных рецепторов; вклад NMDA-рецепторов незначителен. При длительной деполяризации мембраны (ритмическая стимуляция) магниевый блок удаляется и NMDA-каналы начинают проводить ионы Са++, Na+ и К+. Ионы Са+4 через вторичные посредники могут потенцировать (усиливать) минПСП, что может привести, например, к длительному увеличению синаптической проводимости, сохраняющейся часами и даже сутками.
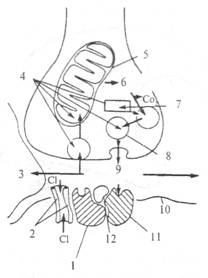
Рис. 2.17. Ответ постсинаптической мембраны на активацию ее глута-матом (схема). а — при небольшой и 6 — при высокой частоте синаптической активации. В случае а глутамат (GLU) активирует как NMDA, так и квисгулатные/каинатные (Q/K) рецепторы, открываются каналы, пропускающие ионы Na+ и К+. NMDA-каналы заблокированы Mg++. В случае б имеет место устойчивая деполяризация постсинаптической мембраны, ионы Mg++ покидают NMDA-каналы и они начинают пропускать ионы Ca++, Na+ и К+. Деполяризация может также активировать вольтзависимые кальциевые каналы. Из тормозных медиаторов ГАМК является самой распространенной в ЦНС. Она синтезируется из L-глутаминовой кислоты в одну стадию ферментом декарбоксилазой, наличие которой является лимитирующим фактором этого медиатора. Известно два типа ГАМК-рецепторов на постсинаптической мембране: ГАМКА (открывает каналы для ионов хлора) и ГАМКБ (открывает в зависимости от типа клетки каналы для К+ или Са++). На рис. 2.18 показана схема ГАМК-рецептора. Интересно, что в его состав входит бензодиазипиновый рецептор, наличием которого объясняют действие так называемых малых (дневных) транквилизаторов (седуксена, тазепама и др.). Прекращение действия медиатора в ГАМК-синапсах происходит по принципу обратного всасывания (молекулы медиатора специальным механизмом поглощаются из синаптической щели в цитоплазму нейрона). Из антагонистов ГАМК хорошо известен бикукулин. Он хорошо проходит через гематоэнцефалический барьер, оказывает сильное воздействие на организм даже в
Рис. 2.18. Схема рецептора постсинаптической мембраны к ГАМК вместе с бензодиазипиновым рецептором. Активация бензодиази-пинового рецептора открывает хлорный канал. 1 — бензодиазипиновый рецептор; 2 — хлорный канал; 3 — глиальная клетка; 4, 9 — ГАМК; 5 — митохондрия; 6 — глютаминовая кислота; 7 — глютаматдекарбоксилаза; 8 — синаптический пузырек; 10 — субсинаптическая мембрана; 11 — ГАМК-рецептор; 12 — ГАМК-модулин.
малых дозах, вызывая конвульсии и смерть. ГАМК обнаруживается в ряде нейронов мозжечка (в клетках Пуркинье, клетках Гольджи, корзинчатых клетках), гиппокампа (в корзинчатых клетках), в обонятельной луковице и черной субстанции. Идентификация ГАМК-цепей мозга трудна, так как ГАМК — обычный участник метаболизма в ряде тканей организма. Метаболическая ГАМК не используется как медиатор, хотя в химическом отношении их молекулы одинаковы. ГАМК определяется по ферменту декарбоксилазе. Метод основан на получении у животных антител к декарбоксилазе (антитела экстрагируют, метят и вводят в мозг, где они связываются с декарбоксилазой). Другим известным тормозным медиатором является глицин. Глицинергические нейроны находятся главным образом в спинном и продолговатом мозге. Считают, что эти клетки выполняют роль тормозных интернейронов.
Рис. 2.19. Ацетилхолинергические пути в головном мозге (схема). 1 — гиппокамп; 2 — базальное ядро; 3 — фронтальная кора; 4 — теменная кора; 5 — затылочная кора.
Ацетилхолин — один из первых изученных медиаторов. Он чрезвычайно широко распространен в периферической нервной системе. Примером могут служить мотонейроны спинного мозга и нейроны ядер черепных нервов. Как правило, холинергические цепи в мозге определяют по присутствию фермента холинэстеразы. В головном мозге тела холинергических нейронов находятся в ядре перегородки, ядре диагонального пучка (Брока) и базальных ядрах. Нейроанатомы считают, что эти группы нейронов формируют фактически одну популяцию холинергических нейронов: ядро переднего мозга, nucleus basalis (оно расположено в базальной части переднего мозга) (рис. 2.19). Аксоны соответствующих нейронов проецируются к структурам переднего мозга, особенно в новую кору и гиппокамп. Здесь встречаются оба типа ацетилхолиновых рецепторов (мускариновые и никотиновые), хотя считается, что мускариновые рецепторы доминируют в более рострально расположенных мозговых структурах. По данным последних лет складывается впечатление, что ацетилхолиновая система играет большую роль в процессах, связанных с высшими интегративными функциями, которые требуют участия памяти. Например, показано, что в мозге больных, умерших от болезни Альцгеймера, наблюдается массивная утрата холинергических нейронов в nucleus basalis. |
||
|
|
Последнее изменение этой страницы: 2018-04-12; просмотров: 460. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |