|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
Анатомия промежуточного мозгаК ЗАДНЕМУ МОЗГУ относятся мозговой мост и мозжечок: Он развивается из четвертого мозгового пузыря. Мостснизу граничит с продолговатым мозгом, сверху переходит в ножки мозга, боковые его отделы образуют средние ножки мозжечка. В передней (вентральной) части моста располагаются скопления серого вещества - собственные ядра моста, в задней (дорсальной) его части лежат ядра верхней оливы, ретикулярной формации и ядра V - VIII пар черепных нервов. Эти нервы выходят на основании мозга сбоку от моста и позади него на границе с мозжечком и продолговатым мозгом. Белое вещество моста в его передней части (основании) представлено поперечно идущими волокнами, направляющимися в средние ножки мозжечка. Они пронизываются мощными продольными пучками волокон пирамидных путей, образующих затем пирамиды продолговатого мозга и направляющихся в спинной мозг. В задней части (покрышке) проходят восходящие и нисходящие системы волокон. Мозжечок (cerebellum) залегает под затылочными долями полушарий большого мозга, отделяясь от него горизонтальной щелью (fissura horizontalis) и располагаясь в задней черепной ямке (fossa cranii posterior). Кпереди от него находится мост и продолговатый мозг. Мозжечок состоит из двух полушарий (hemispheria cerebelli), в каждом из которых выделяют верхнюю (fasies superior) и нижнюю (fasies inferior) поверхности. Кроме того, в мозжечке имеется средняя часть — червь (vermis), отделяющая полушария друг от друга. Серое вещество коры мозжечка (cortex cerebelli), состоящей из тел нейронов, глубокими бороздами делится на дольки. Более мелкие борозды отделяют друг от друга листки мозжечка (folia cerebelli). Кора мозжечка разветвляется и проникает в белое вещество, являющееся телом мозжечка (corpus medullare), образованным отростками нервных клеток. Белое вещество, разветвляясь, проникает в извилины в виде белых пластинок (laminae albae).  Серое вещество содержит парные ядра, залегающие в глубине мозжечка и образующие ядро шатра (nucleus fastigii), относящееся к вестибулярному аппарату. Латеральнее шатра располагаются шаровидное (nucleus globosus) и пробковидное (nucleus emboliformis) ядра, отвечающие за работу мышц туловища, затем зубчатое ядро (nucleus dentalis), контролирующее работу конечностей. Мозжечок связывается с периферией посредством других отделов головного мозга, с которыми он соединяется тремя парами ножек. Верхние ножки (pedunculus cerebellsris superior) соединяют мозжечок со средним мозгом, средние (pedunculus cerebellsris medius) — с мостом, а нижние (pedunculus cerebellsris inferior) — с продолговатым мозгом. Основная функция мозжечка — координация движений, однако, помимо этого, он выполняет некоторые вегетативные функции, принимая участие в управлении деятельностью вегетативных органов и отчасти контролируя скелетную мускулатуру. ФИЗИОЛОГИЯ. Мозжечок не имеет прямой связи с рецепторами организма. Многочисленными путями он связан со всеми отделами центральной нервной системы. К нему направляются афферентные (чувствительные) проводящие пути, несущие импульсы от проприорецепторов мышц, сухожилий, связок, вестибулярных ядер продолговатого мозга, подкорковых ядер и коры больших полушарий. В свою очередь мозжечок посылает импульсы ко всем отделам центральной нервной системы. Функции мозжечка исследуют путем его раздражения, частичного или полного удаления и изучения биоэлектрических явлений. Последствия удаления мозжечка и выпадения его функции итальянский физиолог Лючиани охарактеризовал знаменитой триадой А - астазия, атония и астения. Последующие исследователи добавили еще один симптом - атаксия. Безмозжечковая собака стоит на широко расставленных лапах, совершая непрерывные качательные движения (астазия). У нее нарушено правильное распределение тонуса мышц сгибателей и разгибателей (атония). Движения плохо координированы размашисты, несоразмерны, резки. При ходьбе лапы забрасываются за среднюю линию (атаксия), чего не бывает у нормальных животных. Атаксия объясняется тем, что нарушается контроль движений. Выпадает и анализ сигналов от проприорецепторов мышц и сухожилий. Собака не может попасть мордой в миску с едой. Наклон головы вниз или в сторону вызывает сильное противоположное движение. Движения очень утомляют, животное, пройдя несколько шагов, ложится и отдыхает. Этот симптом называется астенией. С течением времени двигательные расстройства у безмозжечковой собаки сглаживаются. Она самостоятельно ест, походка ее почти нормальна. Только предвзятое наблюдение выявляет некоторые нарушения (фаза компенсации). Как показал Э.А. Асратян, компенсация функций происходит за счет коры головного мозга. Если у такой собаки удалить кору, то все нарушения выявляются снова и уже никогда не компенсируются. Мозжечок участвует в. регуляции движений, делая их плавными, точными, соразмерными. Как показали исследования Л. А. Орбели, у безмозжечковых собак нарушаются вегетативные функции. Константы крови, сосудистый тонус, работа пищеварительного тракта и другие вегетативные функции становится очень неустойчивыми, легко сдвигаются под влиянием тех или иных причин (приём пищи, мышечная работа, изменение температуры и др.). При удалении половины мозжечка нарушения двигательных функций наступают на стороне операции. Это объясняется тем, что проводящие пути мозжечка либо не перекрещиваются вовсе, либо перекрещиваются 2 раза. СРЕДНИЙ МОЗГ На нижней поверхности головного мозга хорошо различимы структуры среднего мозга (mesencephalon): ножки мозга и волокна глазодвигательного нерва (III пара). Первые направляются от переднего края моста, вторые выходят из межножковой ямки и направляются к исчерченным мышцам глазного яблока. Задняя часть среднего мозга называется четверохолмием или пластинкой крыши среднего мозга (lamina tecti mesencephali). Она различима только при удалении мозжечка и затылочных долей полушарий большого мозга. Поперечной бороздой холмики разделяются на верхние и нижние. В двух верхних холмах располагаются подкорковые центры зрения, в нижних — подкорковые центры слуха. На поперечном разрезе виден подкорковый двигательный центр, названный черным веществом (substantia nigra), так как тела его клеток содержат пигмент. Черное вещество имеет полулунную форму и разделяет ножки мозга на вентральную часть — основание и дорсальную — покрышку (tegmentum). В основании находятся волокна, связывающие кору полушарий большого мозга с мозжечком, а также через него проходят проводники сознательных двигательных импульсов. В покрышке содержатся нейроны ретикулярной формации. Покрышка отделяется от крыши среднего мозга (tectum mesencephali) полостью среднего мозга, которая называется водопроводом среднего мозга (aqueductus mesencephali).
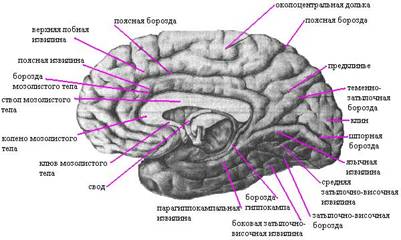
В водопроводе находятся ядра глазодвигательного (III пара) (рис. 264) и блокового (IV пара) нервов. Вокруг водопровода располагается центральное серое вещество, отвечающее за вегетативные функции. Над черным веществом, в латеральном отделе покрышки, залегают проводники суставно-мышечного чувства, сознательной тактильной, температурной и болевой чувствительности, являющиеся волокнами медиальной петли (lemniscus medialis). Латеральная петля (lemniscus lateralis), образованная слуховыми волокнами, располагается дорсальнее. В самой покрышке находится подкорковый центр, получивший название красного ядра (nucleus ruber). Красное ядро отвечает за двигательную автоматизированную деятельность —такую, как ходьба, бег и др. ФИЗИОЛОГИЯ. Средний мозг играет важную роль в регуляции мышечного тонуса и в осуществлении установочных и выпрямительных рефлексов, благодаря которым возможны стояние и ходьба. Роль среднего мозга в регуляции мышечного тонуса лучше всего наблюдать на кошке, у которой сделан поперечный разрез между продолговатым и средним мозгом. У такой кошки резко повышается тонус, мышц, особенно разгибателей. Голова запрокидывается назад, резко выпрямляются лапы. Мышцы настолько сильно сокращены, что попытка согнуть конечность заканчивается неудачей - она сейчас же распрямляется. Животное, поставленное на вытянутые, как палки, лапы, может стоять. Такое состояние называется децеребрационной ригидностью. Если разрез сделать выше среднего мозга, то децеребрационная ригидность не возникает. Примерно через 2 часа такая кошка делает усилие подняться. Сначала она поднимает голову, затем туловище, потом встает на лапы и может начать ходить. Следовательно, нервные аппараты регуляции мышечного тонуса и функции стояния и ходьбы находятся в среднем мозге. Явления децеребрационной ригидности объясняют тем, что перерезкой отделяются от продолговатого и спинного мозга красные ядра и ретикулярная формация. Красные ядра не имеют непосредственной связи с рецепторами и эффекторами, но они связаны со всеми отделами центральной нервной системы. К ним подходят нервные волокна от мозжечка, базальных ядер, коры полушарий большого мозга. От красных ядер начинается нисходящий руброспинальный тракт, по которому передаются импульсы к двигательным нейронам спинного мозга. Его называют экстрапирамидным трактом. Чувствительные ядра среднего мозга выполняют ряд важнейших рефлекторных функций. Ядра, находящиеся в верхних холмиках, являются первичными зрительными центрами. Они получают импульсы от сетчатки глаза и участвуют в ориентировочном рефлексе, т. е. повороте головы к свету. При этом происходит изменение ширины зрачка и кривизны хрусталика (аккомодация), способствующая ясному видению предмета. Ядра нижних холмиков являются первичными слуховыми центрами. Они участвуют в ориентировочном рефлексе на звук - поворот головы в сторону звука. Внезапные звуковые и световые раздражения вызывают сложную реакцию настораживания, мобилизующую животное на быструю ответную реакцию. Промежуточный мозг Анатомия промежуточного мозга Промежуточный мозг располагается под мозолистым телом и сводом, срастаясь по бокам с полушариями большого мозга. К нему относятся: таламус (зрительные бугры), эпиталамус (надбугорная область), метаталамус (забугорная область) и гипоталамус (подбугорная область). Полостью промежуточного мозга является III желудочек. Таламус представляет собой парные скопления серого вещества, покрытые слоем белого вещества, имеющие яйцевидную форму. Передний отдел его примыкает к межжелудочковому отверстию, задний, расширенный, - к четверохолмию. Латеральная поверхность таламуса срастается с полушариями и граничит с хвостатым ядром и внутренней капсулой. Медиальные поверхности обр азуют стенки III желудочка. Нижняя продолжается в гипоталамус. В таламусе различают три основные группы ядер: передние, латеральные и медиальные. В латеральных ядрах происходит переключение всех чувствительных путей, направляющихся к коре больших полушарий. В эпиталамусе лежит верхний придаток мозга - эпифиз, или шишковидное тело, подвешенное на двух поводках в углублении между верхними холмиками пластинки крыши. Метаталамус представлен медиальными и латеральными коленчатыми телами, соединенными пучками волокон (ручки холмиков) с верхними (латеральные) и нижними (медиальные) холмиками пластинки крыши. В них лежат ядра, являющиеся рефлекторными центрами зрения и слуха. Гипоталамус располагается вентральнее зрительного бугра и включает в себя собственно подбугорную область и ряд образований, расположенных на основании мозга. Сюда относятся; конечная пластинка, зрительный перекрест, серый бугор, воронка с отходящим от нее нижним придатком мозга - гипофизом и сосцевидные тела. В гипоталамической области расположены ядра (надзрительное, околожелудочковое и др.), содержащие крупные нервные клетки, способные выделять секрет (нейросекрет), поступающий по их аксонам в заднюю долю гипофиза, а затем в кровь. В заднем отделе гипоталамуса лежат ядра, образованные мелкими нервными клетками, которые связаны с передней долей гипофиза особой системой кровеносных сосудов. Третий желудочек расположен по средней линии и представляет собой узкую вертикальную щель. Боковые стенки его образованы зрительными буграми и подбугорной областью, передняя - столбами свода и передней спайкой, нижняя - образованиями гипоталамуса и задняя - ножками мозга и надбугорной областью. Верхняя стенка - крыша III желудочка, - самая тонкая и состоит из мягкой (сосудистой) оболочки мозга, выстланной со стороны полости желудочка эпителиальной пластинкой (эпендимой). Отсюда в полость желудочка вдавливается большое количество кровеносных сосудов: и образуется сосудистое сплетение. Спереди III желудочек сообщается с боковыми желудочками (I и II) межжелудочковыми отверстиями, а сзади переходит в мозговой Рис 5.Ствол головного мозга, вид сверху и сзади . ФИЗИОЛОГИЯ Главными образованиями промежуточного мозга являются таламус (зрительный бугор) и гипоталамус (подбугорная область). Таламус - чувствительное ядро подкорки. Его называют "коллектором чувствительности", так как к нему сходятся афферентные (чувствительные) пути от всех рецепторов, исключая обонятельные. Здесь находится третий нейрон афферентных путей, отростки которого заканчиваются в чувствительных зонах коры. Главной функцией таламуса является интеграция (объединение) всех видов чувствительности. Для анализа внешней среды недостаточно сигналов от отдельных рецепторов. Здесь происходит сопоставление информации, получаемой по различным каналам связи, и оценка ее биологического значения. В зрительном бугре насчитывается 40 пар ядер, которые подразделяются на специфические (на нейронах этих ядер заканчиваются восходящие афферентные пути), неспецифические (ядра ретикулярной формации) и ассоциативные. Через ассоциативные ядра таламус связан со всеми двигательными ядрами подкорки - полосатым телом, бледным шаром, гипоталамусом и с ядрами среднего и продолговатого мозга. Изучение функций зрительного бугра проводится путем перерезок, раздражения и разрушения. Кошка, у которой разрез сделан выше промежуточного мозга, резко отличается от кошки, у которой высшим отделом центральной нервной системы является средний мозг. Она не только поднимается и ходит, т. е. выполняет сложно координированные движения, но еще проявляет все признаки эмоциональных реакций. Легкое прикосновение вызывает злобную реакцию. Кошка бьет хвостом, скалит зубы, рычит, кусается, выпускает когти. У человека зрительный бугор играет существенную роль в эмоциональном поведении, характеризующемся своеобразной мимикой, жестами и сдвигами функций внутренних органов. При эмоциональных реакциях повышается давление, учащаются пульс, дыхание, расширяются зрачки. Мимическая реакция человека является врожденной. Если пощекотать нос плода 5 - 6 мес., то можно видеть типичную гримасу неудовольствия (П. К. Анохин). При раздражении зрительного бугра у животных возникают двигательные и болевые реакции - визг, ворчание. Эффект можно объяснить тем, что импульсы от зрительных бугров легко переходят на связанные с ними двигательные ядра подкорки. В клинике симптомами поражения зрительных бугров являются сильная головная боль, расстройства сна, нарушения чувствительности как в сторону повышения, так и понижения, нарушения движений, их точности, соразмерности, возникновение насильственных непроизвольных движений. Гипоталамусявляется высшим подкорковым центром вегетативной нервной системы. В этой области расположены центры, регулирующие все вегетативные функции, обеспечивающие постоянство внутренней среды организма, а также регулирующие жировой, белковый, углеводный и водно-солевой обмен. В деятельности вегетативной нервной системы гипоталамус играет такую же важную роль, какую играют красные ядра среднего мозга в регуляции скелетно-моторных функций соматической нервной системы. Самые ранние исследования функций гипоталамуса принадлежат - Клоду Бернару. Он обнаружил, что укол в промежуточный мозг кролика вызывает повышение температуры тела почти на 3°С. Этот классический опыт, открывший локализацию центра терморегуляции в гипоталамусе, получил название теплового укола. После разрушения гипоталамуса животное становится пойкилотермным, т. е. теряет способность удерживать постоянство температуры тела. В холодной комнате температура тела понижается, а в жаркой повышается. Позднее было установлено, что почти все органы, иннервируемые вегетативной нервной системой, могут быть активированы раздражением подбугорной области. Иными словами, все эффекты, которые можно получить при раздражении симпатических и парасимпатических нервов, получаются при раздражении гипоталамуса. В настоящее время для раздражения различных структур мозга широко применяется метод вживления электродов. С помощью особой, так называемой стереотаксической техники, через трепанационное отверстие в черепе вводят электроды в любой заданный участок мозга. Электроды изолированы на всем протяжении, свободен только их кончик. Включая электроды в цепь, можно узко локально раздражать те или иные зоны. При раздражении передних отделов гипоталамуса возникают парасимпатические эффекты - усиление движений кишечника, отделение пищеварительных соков, замедление сокращений сердца и др. при раздражении задних отделов наблюдаются симпатические эффекты - учащение сердцебиения, сужение сосудов, повышение температуры тела и др. Следовательно, в передних отделах подбугорной области располагаются парасимпатические центры, а в задних - симпатические. Так как раздражение при помощи вживленных электродов производится на целом животном, без наркоза, то представляется возможность судить о поведении животного. В опытах Андерсена на козе с вживленными электродами был найден центр, раздражение которого вызывает неутолимую жажду, - центр жажды. При его раздражении коза могла выпивать до 10 л воды. Раздражением других участков можно было заставить сытое животное есть (центр голода). Широкую известность получили опыты испанского ученого Дельгадо на быке с электродом, вживленным в центр страха: Когда на арене разъяренный бык бросался на тореадора, включали раздражение, и бык отступал с ясно выраженными признаками страха. Американский исследователь Д. Олдз предложил модифицировать метод - предоставить возможность самому животному замыкать, что неприятных раздражений животное будет избегать и, наоборот, стремиться повторять приятные. Опыты показали, что имеются структуры, раздражение которых вызывает безудержное стремление к повторению. Крысы доводили себя до истощения, нажимая на рычаг до 14000 раз! Кроме того, обнаружены структуры, раздражение которых, по-видимому, вызывает крайне неприятное ощущение, так как крыса второй раз избегает нажать на рычаг и убегает от него. Первый центр, очевидно, является центром удовольствия, второй - центром неудовольствия. Чрезвычайно важным для понимания функций гипоталамуса явилось открытие в этом отделе мозга рецепторов, улавливающих изменения температуры крови (терморецепторы), осмотического давления (осморецепторы) и состава крови (глюкорецепторы). С рецепторов, обращенных в кровь, возникают рефлексы, направленные на поддержание постоянства внутренней среды организма - гомеостаза. "Голодная кровь", раздражая глюкорецепторы, возбуждает пищевой центр: возникают пищевые реакции, направленные на поиск и поедание пищи. Одним из частых проявлений заболевания гипоталамуса в клинике является нарушение водно-солевого обмена, проявляющееся в выделении большого количества мочи с низкой плотностью. Заболевание носит название несахарного мочеизнурения. Подбугорная область тесно связана с деятельностью гипофиза. В крупных нейронах надзрительного и околожелудочкового ядер гипоталамуса образуются гормоны - вазопрессин и окситоцин. По аксонам гормоны стекают к гипофизу, где накапливаются, а затем поступают в кровь. Иное взаимоотношение между гипоталамусом и передней долей гипофиза. Сосуды, окружающие ядра гипоталамуса, объединяются в систему вен, которые спускаются к передней доле гипофиза и здесь распадаются на капилляры. С кровью к гипофизу поступают вещества - релизинг-факторы, или освобождающие факторы, стимулирующие образование гормонов в передней его доле. Ретикулярная формация. В стволе мозга - продолговатом, среднем и промежуточном мозге, между его специфическими ядрами находятся скопления нейронов с многочисленными сильно ветвящимися отростками, образующими густую сеть. Эта система нейронов получила название сетчатого образования, или ретикулярной формации. Специальные исследования показали, что все так называемые специфические пути, проводящие определенные виды чувствительности от рецепторов к чувствительным зонам коры головного Мозга, дают в стволе мозга ответвления, заканчивающиеся на клетках ретикулярной формации. Потоки импульсов с периферии от экстеро-, интеро- и проприорецепторов. поддерживают постоянное тоническое возбуждение структур ретикулярной формации. От нейронов ретикулярной формации начинаются неспецифические пути. Они поднимаются вверх к коре головного мозга и подкорковым ядрам и спускаются вниз к нейронам спинного мозга. В чем же состоит функциональное значение этой своеобразной системы, не имеющей своей территории, располагающейся между специфическими соматическими и вегетативными ядрами ствола мозга? Методом раздражения отдельных структур ретикулярной формации удалось раскрыть ее функцию как регулятора функционального состояния спинного и головного мозга, а также важнейшего регулятора мышечного тонуса. Роль ретикулярной формации в деятельности центральной нервной системы сравнивают с ролью регулятора в телевизоре. Не давая изображения, он может менять громкость звука и освещенность. Раздражение ретикулярной формации, не вызывая двигательного эффекта, изменяет имеющуюся деятельность, тормозя ее или усиливая. Если у кошки короткими, ритмическими раздражениями чувствительного нерва вызывать защитный рефлекс - сгибание задней лапки, а затем на этом фоне присоединить раздражение ретикулярной формации, то в зависимости от зоны раздражения эффект будет различен: спинальные рефлексы либо резко усилятся, либо ослабятся и исчезнут, т. е. затормозятся. Торможение возникает при раздражении задних отделов ствола мозга, а усиление рефлексов - при раздражении передних отделов. Соответствующие зоны ретикулярной формации получили название тормозящей и активирующей зон. На кору головного мозга ретикулярная формация оказывает активирующее воздействие, поддерживая состояние бодрствования и концентрируя внимание. Если у спящей кошки с вживленными в промежуточный мозг электродами включать раздражение ретикулярной формации, то кошка просыпается, открывает глаза. На электроэнцефалограмме видно, что исчезают медленные волны, характерные для сна, и появляются быстрые волны, свойственные состоянию бодрствования. Ретикулярная формация оказывает на кору головного мозга восходящее, генерализованное (охватывающее всю кору) активирующее влияние. По выражению И.П. Павлова, "подкорка заряжает кору". В свою очередь кора больших полушарий регулирует активность сетчатого образования. КОНЕЧНЫЙ МОЗГ Анатомия конечного мозга Конечный мозг развивается из переднего мозгового пузыря, состоит из сильно развитых парных частей - правого и левого полушария и соединяющей их срединной части. Полушария разделены продольной щелью, в глубине которой лежит пластинка белого вещества, состоящая из волокон, соединяющих два полушария,- мозолистое тело. Под мозолистым телом находится свод, представляющий собой два изогнутых волокнистых тяжа, которые в средней части соединены между собой, а спереди и сзади расходятся, образуя столбы и ножки свода. Спереди от столбов свода находится передняя спайка. Между передней частью мозолистого тела и сводом натянута тонкая вертикальная пластинка мозговой ткани - прозрачная перегородка. Полушарие образовано серым и белым веществом. В нем различают самую большую часть, покрытую бороздами и извилинами, - плащ, образованный лежащим по поверхности серым веществом - корой полушарий; обонятельный мозг и скопления серого вещества внутри полушарий - базальные ядра. Два последних отдела составляют наиболее старую в эволюционном развитии часть полушария. Полостями конечного мозга являются боковые желудочки. В каждом полушарии различают три поверхности: верхнебоковую (верхнелатеральную) выпуклую соответственно своду черепа, срединную (медиальную) - плоскую, обращенную к такой же поверхности другого полушария, и нижнюю - неправильной формы. Поверхность полушария имеет сложный рисунок, благодаря идущим в различных направлениях бороздам и валикам между ними - извилинам. Величина и форма борозд и извилин подвержены значительным индивидуальным колебаниям. Однако существует несколько постоянных борозд, которые ясно выражены у всех и раньше других появляются в процессе развития зародыша. Ими пользуются для разделения полушарий на большие участки, называемые долями. Каждое полушарие делят на пять долей: лобную, теменную, затылочную, височную и скрытую долю, или островок, расположенный в глубине боковой борозды. Границей между лобной и теменной долями является центральная борозда, между теменной и затылочной - теменно-затылочная. Височная доля отделена от остальных боковой бороздой. На верхнелатеральной поверхности полушария в лобной доле различают предцентральную борозду, отделяющую предцентральную извилину, и две лобные борозды: верхнюю и нижнюю, делящие остальную часть лобной доли на верхнюю, среднюю и нижнюю лобные извилины. В теменной доле проходит постцентральная борозда, отделяющая постцентральную извилину, и внутритеменная, делящая остальную часть теменной доли на верхнюю и нижнюю теменные дольки. В нижней дольке выделяют надкраевую и угловую извилины. В височной доле две параллельно идущие борозды - верхняя и нижняя височные - делят ее на верхнюю, среднюю и нижнюю височные извилины. В области затылочной доли наблюдаются поперечные затылочные борозды и извилины. На медиальной поверхности хорошо видны борозда мозолистого тела и поясная, между которыми находится поясная извилина. Над ней, окружая центральную борозду, лежит парацентральная долька. Между теменной и затылочной долями проходит теменно-затылочная борозда, а позади нее - шпорная борозда. Участок между ними называется клином, а лежащий впереди - преклиньем. В месте перехода на нижнюю (базальную) поверхность полушария лежит медиальная затылочно-височная, или язычная, извилина. На нижней поверхности, отделяя полушарие от ствола мозга, проходит глубокая борозда гиппокампа (борозда морского конька), кнаружи от которой находится парагиппокампальная извилина. Латеральнее она отделена коллатеральной бороздой от боковой затылочно-височной извилины. Островок, расположенный в глубине латеральной (боковой) борозды, также покрыт бороздами и извилинами. Кора полушарий большого мозга представляет собой слой серого вещества толщиной до 4 мм. Она образована слоями нервных клеток и волокон расположенных в определенном порядке. Наиболее типично устроенные участки филогенетически более новой коры состоят из шести слоев клеток, старая и древняя кора имеет меньшее количество слоев и устроена проще. Разные участки коры имеют разное клеточное и волокнистое строение. В связи с этим существует учение о клеточном строении коры (цитоархитектоника) и волокнистом строении (миелоархитектоника) коры полушарии большого мозга. Обонятельный мозг у человека представлен рудиментарными образованиями, хорошо выраженными у животных, и составляет наиболее старые участки коры полушарий. Базальные ядра представляют собой скопления серого вещества внутри полушарий. К ним относится полосатое тело, состоящее из хвостатого и чечевицеобразного ядер, соединенных между собой. Чечевицеобразное ядро делится на две части: скорлупу, расположенную снаружи, и бледный шар, лежащий внутри. Они являются подкорковыми двигательными центрами. Кнаружи от чечевицеобразного ядра расположена тонкая пластинка серого вещества - ограда, в переднем отделе височной доли лежит миндалевидное тело. Между базальными ядрами и зрительным бугром находятся прослойки белого вещества, внутренняя, наружная и самая наружная капсулы. Через внутреннюю капсулу проходят проводящие пути. Боковые желудочки (правый и левый) являются полостями конечного мозга, залегают ниже уровня мозолистого тела в обоих полушариях и сообщаются через межжелудочковые отверстия с III желудочком. Они имеют неправильную форму и состоят из переднего, заднего и нижнего рогов и соединяющей их центральной части. Передний рог лежит в лобной доле, он кзади продолжается в центральную часть, которая соответствует теменной доле. Сзади центральная часть переходит в задний и нижний рога, расположенные в затылочной и височной долях. В нижнем роге расположен валик - гиппокамп (морской конек). С медиальной стороны в центральную часть боковых желудочков впячивается сосудистое сплетение, продолжающееся в нижний рог. Стенки боковых желудочков образованы белым веществом полушарий и хвостатыми ядрами. К центральной части снизу примыкает таламус. Белое вещество полушарий занимает пространство между корой и базальными ядрами. Оно состоит из большого количества нервных волокон, идущих в разных направлениях. Выделяют три системы волокон полушарий: ассоциативные (сочетательные), соединяющие части одного и того же полушария; комиссуральные (спаечные), соединяющие части правого и левого полушарий, к которым относятся в полушариях мозолистое тело, передняя спайка и спайка свода, и проекционные волокна, или проводящие пути, соединяющие полушария с лежащими ниже отделами головного мозга и спинным мозгом. Рис 6.Борозды и извилины левого полушария большого мозга; верхнелатеральная поверхность.
Рис 7.Борозды и извилины правого полушария большого мозга; медиальная и нижняя поверхности.
ФИЗИОЛОГИЯ Конечный мозг, или полушария большого мозга, достигшие своего наивысшего развития у человека, справедливо считается самым сложным и самым удивительным созданием природы. Функции этого отдела центральной нервной системы настолько отличаются от функций ствола и спинного мозга, что они выделяются в особую главу физиологии, называемую высшей нервной деятельностью. Этот термин введен И. П. Павловым. Деятельность нервной системы, направленную на объединение и регуляцию всех органов и систем организма, И. П. Павлов назвал низшей нервной деятельностью. Под высшей нервной деятельностью он понимал поведение, деятельность, направленную на приспособление организма к изменяющимся условиям внешней среды, на уравновешивание с окружающей средой. В поведении животного, в его взаимоотношениях с окружающей средой ведущую роль играет конечный мозг, орган сознания, памяти, а у человека - орган умственной деятельности, мышления. Большие достижения И. П. Павлова в области изучения функций полушарий большого мозга объясняются тем, что он доказал рефлекторную природу деятельности коры и открыл присущий только ей новый, качественно высший тип рефлексов, а именно условные рефлексы. Открыв основной механизм деятельности коры полушарий большого мозга, он тем самым создал плодотворный, прогрессивный метод изучения ее функций - метод условных рефлексов. Как выяснилось в дальнейшем, условные рефлексы есть те элементарные акты, те "кирпичики", из которых строится психическая деятельность, или поведение, человека. Значение полушарий у различных животных до И. П. Павлова изучали путем хирургического удаления их. Результаты удаления полушарий большого мозга птиц и собак показали, что вегетативные функции: кровообращение, дыхание, пищеварение и др., существенно не нарушаются. При тщательном уходе животное живет долго. Нарушается его связь с внешней средой. На непосредственно действующие раздражители - укол булавкой, раздражение слизистой оболочки рта пищей - возникает вполне адекватная реакция: лапа отдергивается, пища проглатывается, т. е. у животного сохраняются врожденные безусловные рефлексы. Безвозвратно утрачиваются все приобретенные реакции поведения, все выработанные в процессе индивидуальной жизни условные рефлексы. Для изучения локализации (места нахождения) функций в коре полушарий большого мозга, или, иными словами, значения отдельных зон коры, применяют различные методы: частичное удаление коры, электрическое и химическое раздражение, запись биотоков мозга и метод условных рефлексов. Метод раздражения позволил установить в коре следующие зоны: двигательные (моторные), чувствительные (сенсорные) и немые, которые теперь называют ассоциативными. Двигательные зоны коры. Движения возникают при раздражении коры в области предцентральной извилины. Электрическое раздражение верхней части извилин вызывает движение мышц ног и туловища, средней - рук, нижней - мышц лица. Величина корковой двигательной зоны пропорциональна не массе мышц, а точности движений. Особенно велика зона, управляющая движениями кисти руки, языком, мимической мускулатурой лица. В V слое коры двигательных зон обнаружены гигантские пирамидные клетки (пирамиды Беца), отростки которых спускаются к двигательным нейронам среднего, продолговатого и спинного мозга, иннервирующим скелетную мускулатуру. Путь от коры к двигательным нейронам носит название пирамидного пути. Это путь произвольных движений. После повреждения моторной зоны произвольные движения не могут осуществляться. Раздражение моторной зоны сопровождается движениями на противоположной половине тела, что объясняется перекрестом пирамидных путей на их пути к двигательным нейронам, иннервирующим мышцы. Сенсорные зоны коры. Экстирпация (искоренение) различных участков коры у животных позволила в общих чертах установить локализации сенсорных (чувствительных) функций. Затылочные доли оказались связанными со зрением, височные - со слухом. Зона коры, куда проецируется данный вид чувствительности, называется первичной проекционной зоной. Кожная чувствительность человека, чувства прикосновения, давления, холода и тепла проецируются в постцентральную извилину. В верхней ее части находится проекция кожной чувствительности ног и туловища, ниже - рук и совсем внизу - головы. Абсолютная величина проекционных зон отдельных участков кожи неодинакова. Так, например, проекция кожи кисти рук занимает в коре большую площадь, чем проекция поверхности туловища. Величина корковой проекции пропорциональна значению данной рецептивной поверхности в поведении. Интересно, что у свиньи особенно велика проекция в кору пятачка. Суставно-мышечная, проприоцептивная, чувствительность проецируется в постцентральную и предцентральную извилины. Зрительная зона коры находится в затылочной доле. При раздражении ее возникают зрительные ощущения - вспышки света; удаление ее приводит к слепоте. Удаление зрительной зоны на одной половине мозга вызывает слепоту на одной половине каждого глаза, так как каждый зрительный нерв делится в области основания мозга на две половины (образует неполный перекрест), одна из них идет к своей половине мозга, а другая - к противоположной. При повреждении наружной поверхности затылочной доли не проекционной, а ассоциативной зрительной зоны зрение сохраняется, но наступает расстройство узнавания (зрительная агнозия). Больной, будучи грамотным, не может прочесть написанное, узнает знакомого человека после того, как тот заговорит. Способность видеть - это врожденное свойство, но способность узнавать предметы вырабатывается в течение жизни. Бывают случаи, когда от рождения слепому возвращают зрение уже в старшем возрасте. Он еще долгое время продолжает ориентироваться в окружающем мире на ощупь. Проходит немало времени, пока он научится узнавать предметы с помощью зрения. Функция слуха обеспечивается точными долями больших полушарий. Раздражение их вызывает простые слуховые ощущения. Удаление обеих слуховых зон вызывает глухоту, а одностороннее удаление понижает остроту слуха. При повреждении участков коры слуховой зоны может наступить слуховая агнозия: человек слышит, но перестает понимать значение слов. Родной язык становится ему так же непонятен, как и чужой, иностранный, ему незнакомый. Заболевание носит название слуховой агнозии. Обонятельная зона коры находится на основании мозга, в области парагиппокампальной извилины. Проекция вкусового анализатора, по-видимому, находится в нижней части постцентральной извилины, куда проецируется чувствительность полости рта и языка. Лимбическая система В конечном мозге располагаются образования (поясная извилина, гиппокамп, миндалевидное тело, область перегородки), составляющие лимбическую систему. Они участвуют в поддержании постоянства внутренней среды организма, регуляции вегетативных функций и формировании эмоций и мотиваций. Эту систему иначе называют "висцеральным мозгом", так как эта часть конечного мозга может рассматриваться как корковое представительство интерорецепторов. Сюда поступает информация от внутренних органов. При раздражении желудка, мочевого пузыря в лимбической коре возникают вызванные потенциалы. Электрическое раздражение различных областей лимбической системы вызывает изменения вегетативных функций: кровяного давления, дыхания, движений пищеварительного тракта, тонуса матки и мочевого пузыря. Разрушение отдельных частей лимбической системы приводит к нарушению поведения: животные могут становиться более спокойными или, напротив, агрессивными, легко дающими реакции ярости, изменяется половое поведение. Лимбическая система имеет широкие связи со всеми областями головного мозга, ретикулярной формацией и гипоталамусом. Она обеспечивает высший корковый контроль всех вегетативных функций (сердечно-сосудистой, дыхательной, пищеварительной, обмена веществ и энергии.
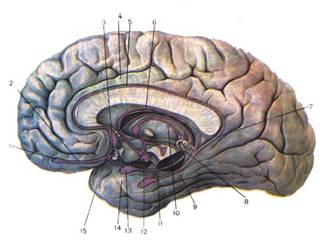
Рис 10.Образования головного мозга, относящиеся к лимбической системе (круг Папеца). 1 - обонятельная луковица; 2 - обонятельный путь; 3 - обонятельный триугольник; 4 - поясная извилина; 5 - серые включения; 6 - свод; 7 - перешейк поясной извилины; 8 - концевая полоска; 9 - гиппокампальная извилина; 11 - гиппокамп; 12 - сосцевидное тело; 13 - миндалевидное тело; 14 - крючок. Ассоциативные зоны коры. Проекционные зоны коры занимают в мозге человека небольшую долю всей поверхности коры. Остальная поверхность занята так называемыми ассоциативными зонами. Нейроны этих областей не связаны ни с органами чувств, ни с мышцами, они осуществляют связь между различными областями коры, интегрируя объединяя все притекающие в кору импульсы в целостные акты научения (чтение, речь, письмо), логического мышления, памяти и обеспечивая возможность целесообразной реакции поведения. При нарушениях ассоциативных зон появляются агнозии - неспособность узнавания и апраксии - неспособность производить заученные движения. Например, стереоагнозия выражается в том, что человек не может найти на ощупь у себя в кармане ни ключа, ни коробки спичек, хотя зрительно он их сейчас же узнает. Выше приводились примеры зрительной агнозии - неспособность прочесть написанное и слуховой - непонимание значения слов. При нарушении ассоциативных зон коры может наступить афазия - потеря речи. Афазия может быть моторной и сенсорной. Моторная афазия возникает при поражении задней трети нижней лобной извилины слева, так называемого центра Брока (этот центр находится только в левом полушарии). Больной понимает речь, но сам говорить не может. При сенсорной афазии, поражении центра Вернике в задней части верхней височной извилины, больной речи не понимает. При аграфии человек разучивается писать, при апраксии - производить заученные движения: зажечь спичку, застегнуть пуговицу, пропеть мелодию и др. Изучение локализации функции методом условных рефлексов на живом здоровом животном позволило И. П. Павлову обнаружить факты, на основе которых им была построена теория динамической локализации функций в коре, затем блестяще подтвержденная при помощи микроэлектродного исследования нейронов. У собак вырабатывали условные рефлексы, например на зрительные раздражения - свет, различные фигуры - круг, треугольник, а затем удаляли всю затылочную, зрительную, зону коры. После этого условные рефлексы исчезали, но проходило время, и нарушенная функция частично восстанавливалась. Это явление компенсации, или восстановления, функции И. П. Павлов объяснил, высказав мысль о существовании ядра анализатора, расположенного в определенной зоне коры, и рассеянных клеток, разбросанных по всей коре, в зонах других анализаторов. За счет этих сохранившихся рассеянных элементов происходит восстановление утраченной функции. Собака может отличать свет от тьмы, но тонкий анализ, установление различий между кругом и треугольником, ей недоступен, он свойствен только ядру анализатора. Микроэлектродное отведение потенциалов от отдельных нейронов коры подтвердило наличие рассеянных элементов. Так, в двигательной зоне коры обнаружили клетки, дающие разряд импульсов на зрительные, слуховые, кожные раздражения, а в зрительной зоне коры выявлены нейроны, отвечающие электрическими разрядами на осязательные, звуковые, вестибулярные и обонятельные раздражители. Кроме того, были найдены нейроны, которые отвечают не только на "свой" раздражитель, как теперь говорят, раздражитель своей модальности, своего качества, но и на один - два чужих. Их назвали полисенсорными нейронами. Динамическая локализация, т. е. способность одних зон замещаться другими, обеспечивает коре высокую надежность.
|
||||||
|
|
Последнее изменение этой страницы: 2018-04-12; просмотров: 331. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |
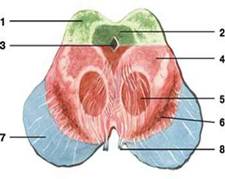
 водопровод.
водопровод.