|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
Вопрос №13. Транспортная роль белков плазмолеммы.Клеточная мембрана- поверхностная, периферическая структура, окружающая протоплазму растительных и животных клеток. Служит не только механическим барьером, ограничивает свободный двусторонний поток в клетку и из нее низко- и высокомолекулярных веществ. Плазмалемма выступает как структура, «узнающая» различные хим., в-ва и регулирующая избирательный транспорт в-в в клетку. Она возникает и обновляется за счет синтетической активности ЭПС и имеет сходное с ними строении. Барьерно-транспортная роль: Механическая устойчивость плазматической мембраны определяется не только свойствами самой мембраны, но и свойствами прилежащих к ней гликокаликса и кортикального слоя цитоплазмы. Внешняя поверхность плазматической мембраны покрыта рыхлым волокнистым слоем вещества толщиной 3-4 нм — гликокаликсом. Он состоит из ветвящихся полисахаридных цепей мембранных интегральных белков, между которыми могут располагаться выделенные клеткой гликолипиды и протеогликаны. Тут же обнаруживаются некоторые клеточные гидролитические ферменты, участвующие во внеклеточном расщеплении веществ (внеклеточное пищеварение, например, в эпителии кишечника). Кортикальный слой цитоплазмы, толщиной 0,1-0,5 мкм, не содержит рибосом и мембранных структур, но богат актиновыми микрофиламентами. Строение плазматической мембраны Плазматическая мембрана, как и другие липопротеидные мембраны клетки, является полупроницаемой. Максимальной проникающей способностью обладает вода и растворенные в ней газы. Транспорт ионов может проходить по градиенту концентраций, т. е. пассивно, без затрат энергии. В этом случае некоторые мембранные транспортные белки образуют молекулярные комплексы, каналы, через которые ионы проходят сквозь мембрану за счет простой диффузии. В других случаях специальные мембранные белки-переносчики избирательно связываются с тем или иным ионом и переносят его через мембрану. Такой тип переноса называется активным транспортом и осуществляется с помощью белковых ионных насосов. Например, затрачивая 1 молекулу АТФ, система К-Nа насоса откачивает за один цикл из клетки 3 иона Nа и закачивает 2 иона К против градиента концентрации. В сочетании с активным транспортом ионов через плазмалемму проникают различные сахара, нуклеотиды и аминокислоты. Макромолекулы, такие как, например, белки, через мембрану не проходят. Они, а также более крупные частицы вещества транспортируются внутрь клетки посредством эндоцитоза. При эндоцитозе определенный участок плазмалеммы захватывает, обволакивает внеклеточный материал, заключает его в мембранную вакуоль. Эта вакуоль — эндосома — сливается в цитоплазме с первичной лизосомой и происходит переваривание захваченного материала. Эндоцитоз формально разделяют на фагоцитоз (поглощение клеткой крупных частиц) и пиноцитоз (поглощение растворов). Плазматическая мембрана принимает участие и в выведении веществ из клетки с помощью экзоцитоза — процесса, обратного эндоцитозу.  Рецепторная роль плазмалеммы Белки-переносчики внешней мембраны клетки являются также рецепторами, узнающими определенные ионы и взаимодействующими с ними. В качестве рецепторов на поверхности клетки могут выступать белки мембраны или элементы гликокаликса. Такие чувствительные к отдельным веществам участки разбросаны по поверхности клетки или собраны в небольшие зоны. Роль многих клеточных рецепторов заключается не только в связывании специфических веществ, но и в передаче сигналов с поверхности внутрь клетки. Например, при действии гормона на клетку цепь событий развертывается следующим образом: молекула гормона специфически взаимодействует с рецепторным белком плазмалеммы и, не проникая в клетку, активирует фермент, синтезирующий ЦАМФ. Последний активирует или ингибирует внутриклеточный фермент или группу ферментов. Разнообразие и специфичность наборов рецепторов на поверхности клеток приводит к созданию очень сложной системы маркеров, позволяющих клеткам отличать «своих» (той же особи или того же вида) от «чужих». Межклеточные соединения У многоклеточных организмов за счет межклеточных взаимодействий образуются сложные клеточные ансамбли. При тесном соседстве клеток друг с другом гликокаликс обеспечивает слипание клеток за счет присутствия в нем трансмембранных гликопротеидов кадгеринов. Это простой межклеточный контакт, при котором зазор между клетками составляет 10-20 нм. В эпителиях часто встречается плотное, или запирающее, соединение, при котором внешние слои двух плазматических мембран максимально сближены и в точках их соприкосновения лежат глобулы интегральных белков мембраны. Такой контакт непроницаем для молекул и ионов, он запирает межклеточные полости. Заякоривающие соединения, или контакты, не только соединяют плазматические мембраны соседних клеток, но и связываются с фибриллярными элементами цитоскелета. Например, для десмосом, имеющих вид бляшек или кнопок, в межклеточном пространстве характерно наличие плотного слоя гликопротеидов десмоглеинов. С цитоплазматической стороны к плазмалемме прилежит слой белка десмоплакина, связанный с промежуточными филаментами цитоскелета. Щелевые контакты считаются коммуникационными соединениями клеток. В зоне щелевого контакта может быть от 20-30 до нескольких тысяч коннексонов — цилидрических белковых структур с внутренним каналом диаметром 2 нм. Каждый коннексон состоит из 6 субъединиц белка коннектина. Коннексоны играют роль прямых межклеточных каналов, по которым ионы и низкомолекулярные вещества могут диффундировать из клетки в клетку.
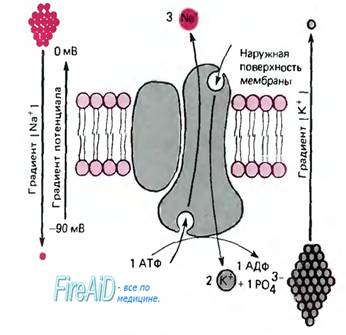
Вопрос №14. Активный транспорт веществ через мембрану. К+ -Na+ насос. Активный транспорт, натриевый насос. В предыдущем разделе описаны пассивная диффузия ионов и образующийся вследствие этого мембранный потенциал при заданных внутри- и внеклеточных концентрациях ионов. Однако в результате этого процесса концентрация ионов внутри клетки стабилизируется не автоматически, поскольку мембранный потенциал несколько более электроотрицателен, чем ЕK, и намного - по сравнению с ENa (около +60 мВ). Благодаря диффузии внутриклеточные концентрации ионов, по крайней мере калия и натрия, должны уравниваться с внеклеточными. Стабильность градиента ионов достигается посредством активного транспорта: мембранные белки переносят ионы через мембрану против электрического и (или) концентрационного градиентов, потребляя для этого метаболическую энергию. Наиболее важный процесс активного транспорта - это работа Na/K-насоса, существующего практически во всех клетках; насос выкачивает ионы натрия из клетки, одновременно накачивая ионы калия внутрь клетки. Таким образом обеспечивается низкая внутриклеточная концентрация ионов натрия и высокая-калия (табл. 1.1). Градиент концентрации ионов натрия на мембране имеет специфические функции, связанные с передачей информации в виде электрических импульсов, а также с поддержанием других активных транспортных механизмов и регулирования объема клетки (см. ниже). Поэтому неудивительно, что более 1/3 энергии, потребляемой клеткой, расходуется на Na/К-насос, а в некоторых наиболее активных клетках на его работу расходуется до 70% энергии.
Шпора 8. |
||
|
|
Последнее изменение этой страницы: 2018-04-12; просмотров: 667. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |