|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
Митоз- непрямое деление клетки.
  Мейоз- редукционное деление клетки. Мейоз- это процесс деления половых клеток. Общий ход Типичный мейоз состоит из двух последовательных клеточных делений, которые соответственно называются мейоз I и мейоз II. В первом делении происходит уменьшение числа хромосом в два раза, поэтому первое мейотическое деление называют редукционным, реже – гетеротипным. Во втором делении число хромосом не изменяется; такое деление называют эквационным (уравнивающим), реже –гомеотипным. Выражения «мейоз» и «редукционное деление» часто используют как синонимы. Интерфаза Предмейотическая интерфаза отличается от обычной интерфазы тем, что процесс репликации ДНК не доходит до конца: примерно 0,2...0,4 % ДНК остается неудвоенной. Таким образом, деление клетки начинается на синтетической стадии клеточного цикла. Поэтому мейоз образно называют преждевременным митозом. Однако в целом, можно считать, что в диплоидной клетке (2n) содержание ДНК составляет 4с. При наличии центриолей происходит их удвоение таким образом, что в клетке имеется две диплосомы, каждая из которых содержит пару центриолей. Первое деление мейоза (редукционное деление, или мейоз I) Сущность редукционного деления заключается в уменьшении числа хромосом в два раза: из исходной диплоидной клетки образуется две гаплоидные клетки с двухроматидными хромосомами (в состав каждой хромосомы входит 2 хроматиды). Профаза 1 (профаза первого деления) состоит из ряда стадий: Лептотена (стадия тонких нитей). Хромосомы видны в световой микроскоп в виде клубка тонких нитей. Раннюю лептотену, когда нити хромосом видны еще очень плохо, называют пролептотена. Зиготена (стадия сливающихся нитей). Происходит конъюгация гомологичных хромосом (от лат.conjugatio – соединение, спаривание, временное слияние). Гомологичные хромосомы (или гомологи) – это хромосомы, сходные между собой в морфологическом и генетическом отношении. У нормальных диплоидных организмов гомологичные хромосомы – парные: одну хромосому из пары диплоидный организм получает от матери, а другую – от отца. При конъюгации образуются биваленты. Каждый бивалент – это относительно устойчивый комплекс из одной пары гомологичных хромосом. Гомологи удерживаются друг около друга с помощью белковых синаптонемальных комплексов. Одинсинаптонемальный комплекс может связывать только две хроматиды в одной точке. Количество бивалентов равно гаплоидному числу хромосом. Иначе биваленты называются тетрады, так как в состав каждого бивалента входит 4 хроматиды. Пахитена (стадия толстых нитей). Хромосомы спирализуются, хорошо видна их продольная неоднородность. Завершается репликация ДНК (образуется особая пахитенная ДНК). Завершаетсякроссинговер – перекрест хромосом, в результате которого они обмениваются участками хроматид. Диплотена (стадия двойных нитей). Гомологичные хромосомы в бивалентах отталкиваются друг от друга. Они соединены в отдельных точках, которые называются хиазмы (от древнегреч. буквы χ – «хи»). Диакинез (стадия расхождения бивалентов). Отдельные биваленты располагаются на периферии ядра. Метафаза I (метафаза первого деления) В прометафазе I ядерная оболочка разрушается (фрагментируется). Формируется веретено деления. Далее происходит метакинез – биваленты перемещаются в экваториальную плоскость клетки. Анафаза I (анафаза первого деления) Гомологичные хромосомы, входящие в состав каждого бивалента, разъединяются, и каждая хромосома движется в сторону ближайшего полюса клетки. Разъединения хромосом на хроматиды не происходит. Процесс распределения хромосом по дочерним клеткам называется сегрегация хромосом. Телофаза I (телофаза первого деления) Гомологичные двухроматидные хромосомы полностью расходятся к полюсам клетки. В норме каждая дочерняя клетка получает одну гомологичную хромосому из каждой пары гомологов. Формируются двагаплоидных ядра, которые содержат в два раза меньше хромосом, чем ядро исходной диплоидной клетки. Каждое гаплоидное ядро содержит только один хромосомный набор, то есть каждая хромосома представлена только одним гомологом. Содержание ДНК в дочерних клетках составляет 2с. В большинстве случаев (но не всегда) телофаза I сопровождается цитокинезом. Интеркинез Интеркинез – это короткий промежуток между двумя мейотическими делениями. Отличается от интерфазы тем, что не происходит репликации ДНК, удвоения хромосом и удвоения центриолей: эти процессы произошли в предмейотической интерфазе и, частично, в профазе I. Второе деление мейоза (эквационное деление, или мейоз II) В ходе второго деления мейоза уменьшения числа хромосом не происходит. Сущность эквационногоделения заключается в образовании четырех гаплоидных клеток с однохроматидными хромосомами (в состав каждой хромосомы входит одна хроматида). Профаза II (профаза второго деления) Не отличается существенно от профазы митоза. Хромосомы видны в световой микроскоп в виде тонких нитей. Метафаза II (метафаза второго деления) В каждой из дочерних клеток формируется веретено деления. Хромосомы располагаются в экваториальных плоскостях гаплоидных клеток независимо друг от друга. Эти экваториальные плоскости могут лежать в одной плоскости, могут быть параллельны друг другу или взаимно перпендикулярны. Анафаза II (анафаза второго деления) Хромосомы разделяются на хроматиды (как при митозе). Получившиеся однохроматидные хромосомы в составе анафазных групп перемещаются к полюсам клеток. Телофаза II (телофаза второго деления) Однохроматидные хромосомы полностью переместились к полюсам клетки, формируются ядра. Содержание ДНК в каждой из клеток становится минимальным и составляет 1с.
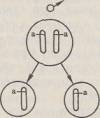
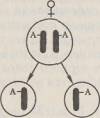
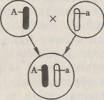
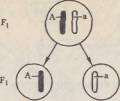
45.Второй закон Менделя. Гипотеза "чистоты гамет" ( на опыте). Закон расщепления, или второй закон Менделя.Если потомков первого поколения, одинаковых по изучаемому признаку, скрестить между собой, то во втором поколении признаки обоих родителей появляются в определенном числовом соотношении: 3/4 особей будут иметь доминантный признак, 'Д — рецессивный. Явление, при котором скрещивание гетерозиготных особей приводит к образованию потомства, часть которого несет доминантный признак, а часть —рецессивный, называется расщеплением. Следовательно, рецессивный признак у гибридов первого поколения не исчез, а был только подавлен и проявится во втором гибридном поколении. Гипотеза чистоты гамет. Мендель предположил, что при образовании гибридов наследственные факторы не смешиваются, а сохраняются в неизменном виде. Вгибриде присутствуют оба фактора — доминантный и рецессивный, но в виде признака проявляется доминантный наследственный фактор, рецессивный же подавляется. Связь между поколениями при половом размножении осуществляется через половые клетки — гаметы. Следовательно, необходимо допустить, что каждая гамета несет только один фактор из пары. Тогда при оплодотворении слияние двух гамет, каждая из которых несет рецессивный наследственный фактор, будет приводить к образованию организма с рецессивным признаком, проявляющимся фенотипически. Слияние же гамет, каждая из которых несет доминантный фактор, или же двух гамет, одна из которых содержит доминантный, а другая рецессивный фактор, будет приводить к развитию организма с доминантным признаком. Таким образом, появление во втором поколении рецессивного признака одного из родителей может быть только при двух условиях: 1) если у гибридов наследственные факторы сохраняются в неизменном виде; 2) если половые клетки содержат только один наследственный фактор из аллельнои пары. Расщепление потомства при скрещивании гетерозиготных особей Мендель объяснил тем, что гаметы генетически чисты, т. е. несут только один ген из аллельнои пары. Гипотезу (теперь ее называют законом) чистоты гамет можно сформулировать следующим образом: при образовании половых клеток в каждую гамету попадает только один ген из аллельнои пары. Известно, что в каждой клетке организма имеется совершенно одинаковый диплоидный набор хромосом. Две гомологичные хромосомы содержат два одинаковых гена. Генетически «чистые» гаметы образуются следующим образом:
При слиянии мужских и женских гамет получается гибрид с диплоидным набором хромосом:
Как видно из схемы, половину хромосом зигота получает от отцовского организма, половину — от материнского. В процессе образования гамет у гибрида гомологичные хромосомы во время I мейотического деления также попадают в разные клетки:
По данной аллельнои паре образуются два сорта гамет. При оплодотворении гаметы, несущие одинаковые или разные аллели, случайно встречаются друг с другом. В силу статистической вероятности при достаточно большом количестве гамет в потомстве 25 % генотипов будут гомозиготными доминантными, 50 % — гетерозиготными, 25 % — гомозиготными рецессивными, т. е. устанавливается отношение 1АА:2Аа:\аа. Соответственно по фенотипу потомство второго поколения при моногибридном скрещивании распределяется в отношении 3:1 (3Д особей с доминантным признаком, '/< особей с рецессивным). Таким образом, при моногибридном скрещивании цитологическая основа расщепления потомства — расхождение гомологичных хромосом и образование гаплоидных половых клеток в мейозе. |
||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2018-04-12; просмотров: 512. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |
||||||||||||||||||||||||||