|
Студопедия КАТЕГОРИИ: АвтоАвтоматизацияАрхитектураАстрономияАудитБиологияБухгалтерияВоенное делоГенетикаГеографияГеологияГосударствоДомЖурналистика и СМИИзобретательствоИностранные языкиИнформатикаИскусствоИсторияКомпьютерыКулинарияКультураЛексикологияЛитератураЛогикаМаркетингМатематикаМашиностроениеМедицинаМенеджментМеталлы и СваркаМеханикаМузыкаНаселениеОбразованиеОхрана безопасности жизниОхрана ТрудаПедагогикаПолитикаПравоПриборостроениеПрограммированиеПроизводствоПромышленностьПсихологияРадиоРегилияСвязьСоциологияСпортСтандартизацияСтроительствоТехнологииТорговляТуризмФизикаФизиологияФилософияФинансыХимияХозяйствоЦеннообразованиеЧерчениеЭкологияЭконометрикаЭкономикаЭлектроникаЮриспунденкция |
Воспроизведение на молекулярном уровне. Репликация ДНК у про- и эукариот.Стр 1 из 17Следующая ⇒ Уровни организации живых систем. Клеточный уровень. Основные положения современной клеточной теории. Молекулярно-генетический уровень( элементарная единица- ген) Клеточный уровень ( клетка) Организменный уровень , по-другому онтогенетический ( особь) Популяционно-видовой ( популяция) Биогеоценотический ( биогеоценозы) Клеточный уровень - это уровень клеток (клеток бактерий, цианобактерий, одноклеточных животных и водорослей, одноклеточных грибов, клеток многоклеточных организмов).элементарные явления представлены реакциями клеточного метаболизма. Благодаря деятельности клетки поступающие извне вещества превращаются в субстраты и энергию ,которые утилизируются в процессе биосинтеза белков в соответствии с существующей информацией . таким образом на клеточном уровне сопрягаются механизмы передачи информации и превращения веществ и энергии. Элементарные явления на этом уровне создают энергетическую и вещественную основу жизни на других уровнях. Клетка - это структурная единица живого, функциональная единица, единица развития. Этот уровень изучают цитология, цитохимия, цитогенетика, микробиология Современная клеточная теория включает следующие основные положения: №1 Клетка - единица строения, жизнедеятельности, роста и развития живых организмов, вне клетки жизни нет;. №2 Клетка - единая система, состоящая из множества закономерно связанных друг с другом элементов, представляющих собой определенное целостное образование; №3 Клетки всех организмов сходны по своему химическому составу, строению и функциям; №4 Новые клетки образуются только в результате деления исходных клеток;  №5 Клетки многоклеточных организмов образуют ткани, из тканей органы. Жизнь организма в целом обусловлена взаимодействием составляющих его клеток; №6 Клетки многоклеточных организмов имеют полный набор генов, но отличаются друг от друга тем, что у них работают различные группы генов, следствием чего является морфологическое и функциональное разнообразие клеток – дифференцировка.
Структурно-функциональная организация про- и эукариотических клеток. Клетки прокариотического типа имеют особенно малые размеры ( не более 0,5-3,0мкм в диаметре) . у них нет морфологически обособленного ядра, т.к. ядерный материал в виде ДНК не отграничен от цитоплазмы оболочкой. В клетке отсутствует развитая система мембран. Генетический аппарат образован единственной кольцевой хромосомой, которая лишена основных белков- гистонов. У прокариот отсутствует клеточный центр. Для них не типичны внутриклеточные перемещения цитоплазмы и амебоидное движение. Время , необходимое для образования двух дочерних клеток ( время генерации), сравнительно мало и исчесляется десятками минут. Прокариотические клетки не делятся митозом. К этому типу клеток относятся бактерии и сине-зеленые водоросли. Эукариотический тип клеточной организации представлен двумя подтипами. Особенностью организмов простейших является то, что они (исключая колониальные формы) соответствуют в структурном отношении уровню одной клетки, а в физиологическом — полноценной особи. В связи с этим одной из черт клеток части простейших является наличие в цитоплазме миниатюрных образований, выполняющих на клеточном уровне функции жизненно важных органов многоклеточного организма. Таковы (например, у инфузорий) цитостом, цитофарингс и порошица, аналогичные пищеварительной системе, и сократительные вакуоли, аналогичные выделительной системе. Клетки многоклеточных организмов имеют оболочку. Плазмолемма ( клеточная оболочка) образована мембраной покрытой снаружи слоем гликокаликса. В клетке выделяют ядро и цитоплазму. В ядре есть оболочка, ядерный сок, ядрышко , хроматин. Цитоплазма представлена основным веществом( матрикс, гиалоплазма), в котором распределены включения и органеллы( шероховатая и гладкая эпс, пластинчатый комплекс, митохондрии, рибосомы, полисомы, лизосомы, периксисомы, микрофибриллы, микротрубочки, центриоли клеточного центра. В растительных клетках выделяют еще и хлоропласты. Жизненный цикл клетки. Его периоды для клеток с разной степенью Дифференцировки. ЖЦК- это период жизни клетки от ее образования ( путем деления материнской клетки) до ее деления или смерти. ЖЦК способных к делению клеток: -Гетерокаталитическая интерфаза -Митотический цикл: -автокаталитическая фаза-подготовка к делению . состоит из G1 периода(синтетический ), S( синтетический ) , G2( постсинтетический). - Митоз. В многоклеточном организме есть клетки которые после своего рождения вступают в период покоя G0 ( это клетки выполняющие специфические функции в составе той или иной функции) ЖЦК не способных к делению клеток: -Гетерокаталитическая интерфаза - Смерть Митотический цикл. Митоз. Биологическое значение митоза. Возможная Патология митоза. Митотический цикл состоит из автокаталитической интерфазы (G1-хромосомы деконденсированные, накапливаются белки и РНК, увеличивается число митохондрий, ;S- репликация ДНК, продолжается синтез белков и РНК;,G2- остановка синтеза ДНК, накапливается энергия, синтезируются РНК и белки, формирующие нити веретена деления) и митоза: Профаза 2n4c – ядерная мембрана растворяется, ядрышко исчезает, происходит конденсация и деспирализация хромосом. Метафаза 2n4c- хромосомы на экваторе клетки. Анафаза 4n4c- хроматиды расходятся к полюсам клетки. Телофаза 2n2c- формирование ядрышка, цитотомия, образование двух дочерних клеток. Биологическое значение митоза. Биологическое значение митоза огромно. Постоянство строения и правильность функционирования органов и тканей многоклеточного организма было бы невозможным без сохранения идентичного набора генетического материала в бесчисленных клеточных поколениях. Митоз обеспечивает важные явления жизнедеятельности, как эмбриональное развитие, рост, восстановление органов и тканей после повреждения, поддержание структурной целостности тканей при постоянной утрате клеток в процессе их функционирования (замещение погибших эритроцитов, слущившихся клеток кожи и прочее). Патологии митоза : Нарушение конденсации хромосом ведет к набуханию и слипанию хромосом Повреждение веретена деления является причиной задержки митоза в метафазе и рассеиванию хромосом Нарушение расхождения хроматид в анафазу митоза ведет к появлению клеток с различным количеством хромосом При отсутствии цитотомии в конце телофазы образуются двух- и многоядерные клетки .
Нуклеиновые кислоты. Строение и функции. Генетический код, его структура и Свойства. ДНК, РНК- макромолекулярные биополимеры , мономерами которых являются нуклеотиды, состоящие из азотистого основания, пентозы( дезоксирибозы или рибозы) и остатка фосфорной кислоты. Соединение азотистого основания и пентозы называется нуклеозидом. Остаток фосфорной к-ты присоединяется к 5’-углероду пентозы. При синтезе ДНК или РНК новый нуклеотид присоединяется 5’- концом к 3’-концу предыдущего. Таким образом полинуклеотидная цепь растет в направлении 5’ ->3’ . У двухцепочечных ДНК полинуклеотидные цепи антипараллельны. Генетический код триплетный. Большинство АК имеют кодоны-синонимы, которые различаются, как правило ,только по третьему нуклеотиду . Свойства ген.кода.: специфичность, вырожденность, непрерывность, универсальность, неперекрываемость, кодон-инициатор, кодоны-терминаторы. Структура : ЦААТ и ТАТА –блоки определяют точность начала транскрипции. Сайленсер-ослабитель , энхансер- усилитель транскрипции. 3’Сайленсер-Энхансер-промотор- зона кэпирования-1й экзон- интрон-последний экзон-сайленсер- энхансер-МГЭ-мини спейсеры 5’
Воспроизведение на молекулярном уровне. Репликация ДНК у про- и эукариот. Одна из основных функций ДНК- сохранение и передача наследственной информации. В основе этой функции лежит способность ДНК к самокопированию- репликация. В результате репликации из одной материнской молекулы ДНК образуются две дочерние молекулы ДНК- копии материнской. Геликаза-расплетает двойную спираль ДНК Дестабилизирующие белки – выпрямляют цепи ДНК ДНК-топоизомераза- разрывает фосфодиэфирные связи в одной из цепей ДНК, снимает напряжение спирали. РНК-праймаза- обеспечивает синтез РНК-затравки для фрагментов Оказаки ДНК-полимеразы- синтез полинуклеотидной цепи в направлении 5-3 ДНК-лигаза –сшивает фрагменты Оказаки после удаления ДНК-затравки. Понятие о репарации ДНК.
Вопрос Ген - это структурная и функциональная единица генома.Геном эукариот организован сложнее, чем у прокариот. Для него характерен хромосомный уровень организации. В хромосомах ДНК находится в окружении белков Гены – это участки ДНК с определенной последовательностью нуклеотидов, кодирующие функциональную молекулу РНК (мРНК, тРНК, рРНК). В состав генов входят регуляторные участки (промоторы), а также последовательности, кодирующие синтез РНК. Гены эукариот имеют прерывистое строение и состоят из экзонов (кодирующие участки) и интронов (некодирующие элементы) 8 вопрос! Экспрессия экспрессия гена -процесс построения матричной РНК по участку молекулы ДНК На первом этапе экспрессии генов происходит переписывание генетической информации, заключенной в генах, на матричные (информационные) РНК (мРНК ), которые являются местом промежуточного хранения этой информации при ее реализации. В некоторых случаях уже сами РНК являются конечным результатом экспрессии генов, и после ряда ферментативных модификаций они непосредственно используются в клеточных процессах. Это относится, прежде всего, к рибосомным и транспортным РНК (рРНК и тРНК), которые вместе составляют основную часть суммарной РНК клетки. К таким РНК принадлежат и малые ядерные РНК (мяРНК), участвующие в процессинге предшественников мРНК эукариот, РНК, входящие в состав ферментов, и природные антисмысловые РНК Транскрипция В процессе транскрипции генов происходит биосинтез молекул РНК, комплементарных одной из цепей матричной ДНК, сопровождаемый полимеризацией четырех рибонуклеозидтрифосфатов (ATP, GTP, CTP и UTP) с образованием 3'-5'-фосфодиэфирных связей и освобождением неорганического пирофосфата. Основными ферментами, осуществляющими транскрипцию, являются ДНК-зависимые РНК-полимеразы, которые функционируют с участием многочисленных факторов транскрипции - регуляторных белков, осуществляющих высокоспецифические белок-белковые и белково-нуклеиновые реакции.Благодаря факторам транскрипции и регуляторным последовательностям генов становится возможным специфический синтез РНК и осуществляется регуляция экспрессии генов на уровне транскрипции. В участке ДНК, соответствующем отдельному гену перед структурной частью, в которой зашифрована последовательность аминокислот в пептиде, обязательно располагается последовательность нуклеотидов, узнаваемая РНК-полимеразой. Такая последовательность называется промотором. РНК-полимераза находит промотор, взаимодействует с ним и после этого, двигаясь вдоль молекулы ДНК, обеспечивает постепенную сборку молекулы иРНК в соответствии с принципом комплементарности и антипараллельности. В конце структурной части гена расположен участок с особой последовательностью нуклеотидов - терминатор. Он обязательно включает один из нонсенс-триплетов, не кодирующих аминокислоты. В результате транскрипции синтезируется молекула преинформационной РНК Процессинг - модифицирование первичного транскрипта и удаление из него некодирующих интронных участков с последующим соединением( сплайсингом) кодирующих последовательностей- экзонов. Основные стадии процессинга: 1. отщепление концевых участков первичного транскрипта (спейсеров); 2. формирование на 5/; конце колпачка, состоящего из особой последовательности нуклеотидов; 3. формирование на 3' конце полиадениловой последовательности нуклеотидов АААА: 4. метилирование некоторых внутренних азотистых оснований в транскрипте, стабилизирующее молекулу РНК; 5. вырезание неинформативных участков, соответствующих нитронам ДНК, и сшивание (сплайсинг) участков, соответствующих экзонам. Вырезание интронов происходит с участием сплайссом. Некодирующие последовательности - интроны превращаются в малую ядерную РНК (мяРНК). Выделено до 30 мяРНК, они участвуют в сплайсинге и ядерноцитоплазматическом транспорте белков. В результате процессинга у эукариот зрелая нРНК характеризуется следующими особенностями строения: Кэп -особая последовательность нуклеотидов с метилированными основаниями, которая обеспечивает узнавание малых субъединиц рибосом; Лидер -вводная последовательность нуклеотидов, комплементарная последовательности в молекуле рРНК малой субъединицы рибосом, которая обеспечивает прикрепление иРНК к малой субъединице. Стартовый кодон - триплет нуклеотидов, кодирующий в большинстве случаев аминокислоту формилметионин (АУГ). Кодирующая часть - последовательность колонов, шифрующих определенную последовательность аминокислот в соответствующей пептидной цепи. Поли А-хвост - концевая часть молекулы иРНК, включающая нонсенс-кодон и поли-А последовательность.
Регуляция экспрессии генов у эукариот. 9 вопрос! Трансляция и пострансляционные процессы Трансляция - процесс сборки пептидной цепи, происходящей в цитоплазме на рибосомах на основании программы, содержащейся в иРНК. Основные фазы трансляции: 1) инициация; 2) элонгация; 3) терминация . Инициация трансляция предполагает следующие события: • с помощью колпачка иРНК находит в цитоплазме малую субъединицу рибосомы; • с помощью лидерной последовательности устанавливается связь с комплементарным участком определенной фракции рРНК и иРНК прикрепляется к малой субъединице; • к стартовому кодону (АУГ) присоединяется тРНК, несущая формилметионин • малая субъединица ассоциируется с большой субъединицей в аминоацильном центре (АЦ), в которой располагается формилметионин. Таким образом, фаза инициация завершается формированием комплекса иРНК и рибосомы с подстановкой начальной для всех пептидных цепей аминокислоты - формилметионин. Фаза элонгации, т.е. наращивание пептидной цепи. Осуществляется путем постепенной подстановки аминокислоты в соответствии с очередным кодовом иРНК, который встает против аминоацильного центра. К этому кодону присоединяется соответствующая тРНК, имеющая комплементарный ему антикодон. Она несет определенную аминокислоту, которая располагается в аминоацильном центре (АЦ), тРНК, соединенная с предыдущим кодовом оказывается в пептидильном центре (ПЦ где располагает свою аминокислоту (цепочку АК). Между двумя аминокислотами, расположенными в пептидильном и аминоацильном центре, при участии имеющихся здесь ферментов возникает пептидная связь: После установления пептидной связи предыдущая тРНК отделяется от своей аминокислоты и своего кодона и уходит в цитоплазму, а последняя тРНК, нагруженная цепочкой аминокислот, переходит в ПЦ, заставляя иРНК перемещаться вдоль рибосомы и устанавливать новый кодон против аминоацильного центра. После прохождения через рибосому всей кодирующей части иРНК на рибосоме собирается пептидная цепь с определенной последовательностью аминокислот. Фаза терминации наступает, когда в контакт с рибосомой приходит концевой участок иРНК, который включает нонсенс-кодон, не кодирующий никакой аминокислоты. На этом сборка пептидной цепи заканчивается. По мере освобождения 5/ конца иРНК колпачок может находить новые малые субьединицы рибосом и процесс трансляции может повторно осуществляться на новых рибосомах. Комплекс рибосом, находящихся в контакте с одной молекулой иРНК и синтезирующих одинаковые пептидные цепи, называется полирибосомой ( полисомой)
Посттрансляционные процессы. В ходе предыдущих этапов реализации наследственной информации обеспечивается синтез пептидной цепи, которая в большинстве случаев начинается с аминокислоты формилметионин и соответствует первичной структуре белковой молекулы. Последующие события заключаются в отщеплении формилметионина, в некоторых случаях осуществляется модифицирование пептида после трансляции, формируется вторичная и третичная структуры белка (иногда для некоторых белков, характеризующихся четвертичной структурой, осуществляется объединение одинаковых, либо различных пептидных цепей с образованием активно функционирующего белка). В зависимости от того, каковы функции белка (ферменты, строительный материал, антитела и т.д), он принимает участие в обеспечении морфо-функциональных особенностей клетки (организма), т.е. в формировании определенных сложных признаков. Это является завершающим этапом процесса реализации генетической информация. У эукариот образование РНК происходит и в цитоплазме: в митохондриях и хлоропластах (у растений), обладающих собственной системой синтеза белка и собственной генетической информацией в виде ДНК - цитоплазматическая наследственность, однако, система белкового синтеза в митохондриях и пластидах аналогична таковой у прокариот и существенно отличается от белкового синтеза в ядре высших животных. Гены расположенные в цитоплазме вне хромосом, называются плазмогенами. Ими объясняется особый тип наследования, при котором признак передается через цитоплазму яйцеклетки (по материнской линии). Уникальной остае 10 вопрос! Хромосомы, хроматин.
Хромосомы - нуклеопротеидные структурные Элементы ядра клетки, содержащие, днк, в которой заключена наследственная Информация организма, способны к самовоспроизведению, обладают структурной и функциональной индивидуальностью и сохраняют её в ряду поколений.
Химическая организация хромосом.
Химический состав хромосом - ДНК- 40%, Гистоновых белков - 40%. Негистоновых - 20% немного РНК. Липиды,полисахариды,ионы металлов.
Имеется 5 фракций основных гистоновых белков (Н1 , Н2А , Н2В , НЗ , Н4) и более 100 фракций кислых негистоновых белков,
Функции гистоновых белков: регуляторная (прочно соединяясь с ДНК препятствуют считыванию информации) и структурная (обеспечивают пространственную организацию ДНК в хромосомах. Образуя Нуклеогистон). Функции негистоновых белков: среди них ферменты регулирующие Процессы: • синтеза РНК (полимеразы) и процессинга РНК, • редупликации и репарации ДНК (геликаза. ДНК Попимераза, эндонуклеаза. Экзонуклеаза, лигаза), • регуляторная функция, заключающаяся в «запрещении» или «разрешении» считыв информации с молекулы ДНК
Днк эукариотических клеток представлена Следующими фракциями: а) уникальные последовательности генов 56 % - Присутствуют в гаплоидном наборе в единственном числе, образуют основную часть Структурных и регуляторных генов, б) гены со средним числом повторов 8 % - 102 - 104 копии это структурные гены кодирующие первичную структуру гистонов или Нуклеотидов рибосомальных и транспортных РНК, в) многократно повторяющиеся гены 12 % г) блуждающие структурные гены, Положение которых в хромосоме меняется в зависимости от жизненного цикла, д) молчащие гены они реплицируются, но не Транскрибируются. Участвуют в обеспечении структурной организации хроматина и В регуляции экспрессии генов.
Классификация и функции хроматина: различают гетеро- и эухроматин. а) гетерохроматин: • факультативный - образуется при Спирализации одной из двух гомологичных хромосом. Типичным примером служит Тельце полового хроматина, образуемого одной из двух Х-хромосом соматических Клеток женских особей человека и млекопитающих Функция Факультативного гетерохроматина заключается в компенсации снижении дозы определенного Гена. • структурный ~ отличается Высокоспирализованным состоянием, которое сохраняется на протяжении всего мит. Цикла. Он занимает постоянные участки в гомологичных хромосомах - это фрагменты Околоцентромерных, теломерных участков хромосом, Не содержит структурных генов (нетранскрибируемый); он выполняет опорную Функцию. б) эухроматин - имеет менее компактную организацию, деспирализуется в Конце митоза, образует слабоокрашенные нитчатые структуры содержит структурные транскрибируемые Гены: в каждой хромосоме свой порядок расположения эухроматина и гетерохроматина. В структурном отношении хроматин представляет собой нитчатые комплексные молекулы дезоксирибонуклеопротеида (ДНП), которые состоят из ДНК, ассоциированной с Гистонами Уровни упаковки ДНК (ДНП) в хромосому (нуклеосомный, хроматиновая фибрилла, интерфазная и метафазная хромосомы). Строение метафазной хромосомы ДНК претерпевает пять уровней компактизацни от 2 нм (ее собственный диаметр) до 1400 нм (высококонденсированная метафазная хромосома). Низшим уровнем иерархической организации хромосом считается нуклеосомный. Нуклеосома состоит из кора (сердцевины, стержня) и намотанной на негоДНК. Кор представляет собой гистоновый октамер. Хроматин на этой стадии имеет вид «бусин» нанизанных на «нить» (молекулярную ДНК). Такая структура обеспечивает компактизацию примерно в 6—7 раз. Вторая ступень компактизации - формирование хроматиновой фибриллы диаметром 30 нм. В этом процессе участвует гистон HI, который связывается с ДНК между нуклеосомными корами и сворачивает нуклеосомную фибриллу в спираль, наполобие соленоида, с шагом в 6-8 нуклеосом. Уровень компактизации на этом этапе достигает примерно 40. Третий этап — петельно-доменный — наиболее сложный. Соленоидная фибрилла складывается 11 вопрос! Кариотип – это совокупность метафазных хромосом, характерных для определенного вида организмов. Постоянство кариотипа поддерживается с помощью точных механизмов митоза и мейоза Кариотип используется в качестве видовой характеристики (существует особый раздел систематики – кариосистематика). Кариотипический критерий является одним из важнейших критериев вида. Сущность этого критерия заключается в том, что все особи данного вида характеризуются определенным кариотипом. В понятие «кариотип» включается число хромосом, их размеры, морфология, особенности продольной дифференцировки. Если оба плеча хромосомы равны по длине, то такая хромосома называется метацентрической, если неравны – то такая хромосома называется субметацентрической, если же одно из плеч очень короткое, то такая хромосома называется акроцентрической. Конечные участки хроматид называются теломеры. У некоторых хромосом в области теломер имеются удаленные структуры (спутники); это спутничные хромосомы. В 1960 г. в г. Денвере (США) была разработана первая классификация хромосом человека. В ее основу легли размеры хромосом и положение первичной перетяжки — центромеры. Все хромосомы по форме разделены на метацентрические, субметацентрические и акроцентрические и подразделены на 7 групп, обозначенных латинскими буквами А, В, С, D, Е, F и G Классификация хромосом человека по размеру и расположению центромера Характеристика хромосом А(1)1,2,3 1 и 3 почти метацентрические и 2-крупная субметацентрическая В (11) 4,5 крупные субакроцентрические С (III) 6-12 средние субметацентрические A(lV) 13-15 средние акроцентрические E(V) 16-18 мелкие субметацентрические F(VI) 19-20 самые мелкие мегацентрические G(VII) 21-22 самые мелкие акроцентрические Х-хромосома (относится к III группе 23 средняя почти метацентрическая Y-хромосома 23 мелкая акроцентрическая
Вопрос 13-14 Cперматогенез
Фазы сперматогенеза: Сперматогенез осуществляется в семенниках и подразделяется на четыре фазы: 1) размножения, 2) роста, 3) созревания, 4) формирования. Во время фазы размножения диплоидные сперматогонии многократно делятся митозом. Часть образовавшихся сперматогониев может подвергаться повторным митотическим делениям, в результате чего образуются такие же клетки сперматогонии. Другая часть прекращает делиться и увеличивается в размерах, вступая в следующую фазу сперматогенеза — фазу роста. Фаза роста соответствует интерфазе 1 мейоза, т.е. во время нее происходит подготовка клеток к мейозу. Главным событием фазы роста является репликация ДНК. Во время фазы созревания клетки делятся мейозом; во время первого деления мейоза они называются сперматоцитами 1-го порядка, во время второго — сперматоцитами 2-го порядка. Из одного сперматоцита 1-го порядка возникают четыре гаплоидные сперматиды. Фаза формирования характеризуется тем, что первично шаровидные сперматиды подвергаются ряду сложных преобразований, в результате которых образуются сперматозоиды. В нем участвуют все элементы ядра и цитоплазмы. У человека сперматогенез начинается в период полового созревания; срок формирования сперматозоида — три месяца, т.е. каждые три месяца сперматозоиды обновляются. Сперматогенез происходит непрерывно и синхронно в миллионах клеток. Строение сперматозоида
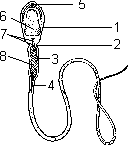
Строение сперматозоида: 1 — «головка»; 2 — «шейка»; 3 — средняя часть; 4 — жгутик; 5 — акросома; 6 — ядро; 7 — центриоли; 8 — митохондрии.Сперматозоид млекопитающих имеет форму длинной нити. Длина сперматозоида человека 50–60 мкм. В строении сперматозоида можно выделить «головку», «шейку», промежуточный отдел и хвостик. В головке находится ядро и акросома. Ядро содержит гаплоидный набор хромосом. Акросома — мембранный органоид, содержащий ферменты, используемые для растворения оболочек яйцеклетки. В шейке расположены две центриоли, в промежуточном отделе — митохондрии. Хвостик представлен одним, у некоторых видов — двумя и более жгутиками. Жгутик является органоидом движения и сходен по строению со жгутиками и ресничками простейших. Для движения жгутиков используется энергия макроэргических связей АТФ, синтез АТФ происходит в митохондриях. Сперматозоид открыт в 1677 году А. Левенгуком. Овогенез Осуществляется в яичниках, подразделяется на три фазы: 1) размножения, 2) роста, 3) созревания.Во время фазы размножения диплоидные овогонии многократно делятся митозом. Фаза роста соответствует интерфазе 1 мейоза, т.е. во время нее происходит подготовка клеток к мейозу: клетки значительно увеличиваются в размерах вследствие накопления питательных веществ. Главным событием фазы роста является репликация ДНК. Во время фазы созревания клетки делятся мейозом. Во время первого деления мейоза они называются овоцитами 1-го порядка. В результате первого мейотического деления возникают две дочерние клетки: мелкая, называемая первым полярным тельцем, и более крупная — овоцит 2-го порядка. Во время второго мейотического деления овоцит 2-го порядка делится с образованием яйцеклетки и второго полярного тельца, а первое полярное тельце — с образованием третьего и четвертого полярных телец. Таким образом, в результате мейоза из одного овоцита 1-го порядка образуются одна яйцеклетка и три полярных тельца.
Фазы овогенеза: В отличие от образования сперматозоидов, которое происходит только после достижения половой зрелости, процесс образования яйцеклеток у человека начинается еще в эмбриональном периоде и течет прерывисто. У зародыша полностью осуществляются фазы размножения и роста и начинается фаза созревания. К моменту рождения девочки в ее яичниках находятся сотни тысяч овоцитов 1-го порядка, остановившихся, «застывших» на стадии диплотены профазы 1 мейоза — первый блок овогенеза. В период полового созревания мейоз возобновится: примерно каждый месяц под действием половых гормонов один из овоцитов (редко два) будет доходить до метафазы 2 мейоза — второй блок овогенеза. Мейоз может пройти до конца только при условии оплодотворения; если оплодотворение не происходит, овоцит 2-го порядка погибает и выводится из организма. Строение яйцеклеток Форма яйцеклеток обычно округлая. Размеры яйцеклеток колеблются в широких пределах — от нескольких десятков микрометров до нескольких сантиметров (яйцеклетка человека — около 120 мкм). К особенностям строения яйцеклеток относятся: наличие оболочек, располагающихся поверх плазматической мембраны и наличие в цитоплазме более или менее большого количества запасных питательных веществ.
Яйцеклетка млекопитающих: У большинства животных яйцеклетки имеют дополнительные оболочки, располагающиеся поверх цитоплазматической мембраны. В зависимости от происхождения различают: первичные, вторичные и третичные оболочки. Первичные оболочки формируются из веществ, выделяемых овоцитом. Образуется слой, контактирующий с цитоплазматической мембраной яйцеклетки. Он выполняет защитную функцию, обеспечивает видовую специфичность проникновения сперматозоида, т.е. не позволяет сперматозоидам других видов проникать в яйцеклетку. У млекопитающих эта оболочка называется блестящей. Вторичные оболочки образуются выделениями фолликулярных клеток яичника, имеются далеко не у всех яйцеклеток. Вторичная оболочка яиц насекомых содержит канал — микропиле, через который сперматозоид проникает в яйцеклетку. Третичные оболочки образуются за счет деятельности специальных желез яйцеводов. Например, из секретов особых желез формируются белковая, подскорлуповая пергаментная, скорлуповая и надскорлуповая оболочки у птиц и рептилий. Вторичные и третичные оболочки образуются у яйцеклеток животных, зародыши которых развиваются во внешней среде. Поскольку у млекопитающих наблюдается внутриутробное развитие, их яйцеклетки имеют только первичную оболочку, поверх которой располагается лучистый венец — слой фолликулярных клеток, доставляющих к яйцеклетке питательные вещества. В яйцеклетках происходит накопление запаса питательных веществ, которые называют желтком. Он содержит жиры, углеводы, РНК, минеральные вещества, белки, причем основную его массу составляют липопротеиды и гликопротеиды. Желток содержится в цитоплазме в виде желточных гранул. Количество питательных веществ, накапливаемых в яйцеклетке, зависит от условий, в которых происходит развитие зародыша. Если развитие яйцеклетки происходит вне организма матери и приводит к формированию крупных животных, то желток может составлять более 95% объема яйцеклетки. Яйцеклетки млекопитающих, развивающиеся внутри тела матери, содержат малое количество желтка — менее 5%, так как питательные вещества, необходимые для развития, эмбрионы получают от матери.
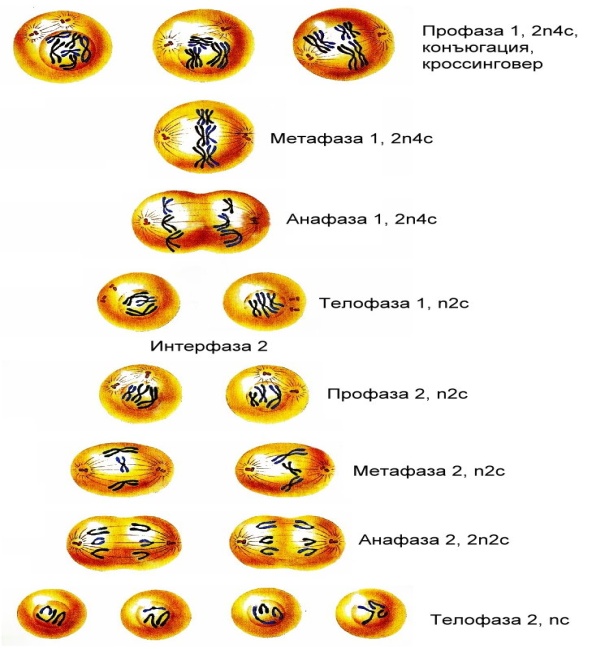
Типы яйцеклеток хордовых животных: В зависимости от количества содержащегося желтка различают следующие типы яйцеклеток: алецитальные (не содержат желтка или имеют незначительное количество желточных включений — млекопитающие, плоские черви); изолецитальные (с равномерно распределенным желтком — ланцетник, морской еж); умеренно телолецитальные (с неравномерно распределенным желтком — рыбы, земноводные); резко телолецитальные (желток занимает большую часть, и лишь небольшой участок цитоплазмы на анимальном полюсе свободен от него — птицы). В связи с накоплением питательных веществ, у яйцеклеток появляется полярность. Противоположные полюсы называются вегетативным и анимальным. Поляризация проявляется в том, что происходит изменение местоположения ядра в клетке (оно смещается в сторону анимального полюса), а также в особенностях распределения цитоплазматических включений (во многих яйцах количество желтка возрастает от анимального к вегетативному полюсу). Яйцеклетка человека была открыта в 1827 году К.М. Бэром. Вопрос15 Мейоз – особый способ деления эукариотических клеток, в результате которого образуются клетки со уменьшенным в два раза набором хромосом, образованные клетки имеют различный набор аллелей генов – генетически неодинаковы, эти клетки превращаются в гаметы (у животных) или споры (у растений и грибов). Мейоз состоит из двух последовательных делений, которым предшествует однократная репликация ДНК. Первое мейотическое деление (мейоз 1) называется редукционным, поскольку именно во время этого деления происходит уменьшение числа хромосом вдвое: из одной диплоидной клетки (2n4c) образуются две гаплоидные (1n2c). Интерфаза 1 (в начале – 2n2c, вконце – 2n4c) происходит обычно и сопровождается ростом, синтезом и накоплением веществ и энергии, необходимых для осуществления обоих делений, увеличением числа органоидов, удвоением центриолей, репликацией ДНК, которая завершается в профазе 1. Профаза 1 (2n4c). Самая продолжительная и сложная фаза мейоза. Состоит из ряда последовательных стадий. Лептотена, стадия тонких нитей. Хромосомы слабо конденсированы. Они уже двухроматидные (каждая хромосома состоит из двух сестринских хроматид), но хроматиды настолько сближены, что хромосомы имеют вид длинных одиночных тонких нитей. Теломеры хромосом еще прикреплены к ядерной мембране с помощью особых структур – прикрепительных дисков. Зиготена, стадия сливающихся нитей. Гомологичные хромосомы начинают притягиваться друг к другу сходными участками и конъюгируют. Конъюгацией называют процесс тесного сближения гомологичных хромосом. (Процесс конъюгации также называют синапсисом.). Начинается распад ядерной оболочки на фрагменты, происходит расхождение центриолей к разным полюсам клетки, формирование нитей веретена деления, «исчезновение» ядрышек, продолжается конденсация двухроматидных хромосом. Происходит процесс, отсутствующий при митозе – конъюгация, процесс тесного сближения и гомологичных хромосом. Пару конъюгирующих гомологичных хромосом называют бивалентом (это пара хромосом), или тетрадой (в биваленте четыре хроматиды). Полагают, что каждый ген приходит в соприкосновение с гомологичным ему геном другой хромосомы, количество бивалентов равно гаплоидному набору хромосом. Пахитена, стадия толстых нитей. Процесс спирализации хромосом продолжается, причем в гомологичных хромосомах он происходит синхронно. Становится хорошо заметно, что хромосомы двухроматидные. В пахитене наблюдается особенно тесный контакт между хроматидами. Важнейшим событием пахитены является кроссинговер – обмен участками между несестринскими хроматидами гомологичных хромосом. Кроссинговер приводит к первой во время мейоза рекомбинации генов. Диплотена. Хромосомы в бивалентах перекручиваются и начинают отталкиваться друг от друга. Процесс отталкивания начинается в области центромеры и распространяется по всей длине бивалентов. Однако они все еще остаются связанными друг с другом в некоторых точках. Их называют хиазмы. Эти точки появляются в местах кроссинговера. В ходе гаметогенеза у человека может образовываться до 50 хиазм.
Диакинез. Хромосомы максимально укорачиваются и утолщаются за счет спирализации хроматид, ядерная оболочка почти полностью разрушена. Происходит сползание хиазм к концам хроматид. Метафаза 1 (2n4c) происходит выстраивание бивалентов в экваториальной плоскости клетки, прикрепление микротрубочек веретена деления одним концом – к центриолям, другим – к центромерам хромосом, а не к центромерам хроматид, как это было при митозе. Анафаза 1 (2n4c) – случайное независимое расхождение двухроматидных хромосом к противоположным полюсам клетки (из каждой пары гомологичных хромосом одна хромосома отходит к одному полюсу, другая – к другому). Происходит вторая рекомбинация генетического материала – у каждого полюса оказывается гаплоидный набор двухроматидных хромосом, часть из них – отцовские, часть – материнские. Многие хроматиды в хромосомах после кроссинговера стали мозаичными, одновременно несут некоторые гены отца и матери. Телофаза 1 (1n2c в каждой клетке). Происходит образование ядерных оболочек вокруг гаплоидных наборов двухроматидных хромосом, деление цитоплазмы. Из одной диплоидной клетки (2n4c) образовались две клетки с гаплоидным набором хромосом (n2c), поэтому это деление называют редукционным. Интерфаза 2, или интеркинез (1n2c) представляет собой перерыв между первым и вторым мейотическими делениями, продолжительность этого периода различается у разных организмов – в некоторых случаях обе дочерние клетки сразу вступают во второе деление, а иногда второе деление начинается через несколько месяцев или лет. Но так как хромосомы двухроматидные, во время интерфазы 2 не происходит репликация ДНК. Второе мейотическое деление (мейоз 2) называется эквационным. Профаза 2 (1n2c). Короче профазы 1, хроматин конденсирован, нет конъюгации и кроссинговера, происходят процессы, обычные для профазы – распад ядерных мембран на фрагменты, расхождение центриолей к разным полюсам клетки, формирование нитей веретена деления. Метафаза 2 (1n2c). Двухроматидные хромосомы выстраиваются в экваториальной плоскости клетки, формируется метафазная пластинка. Создаются предпосылки для третьей рекомбинации генетического материала – многие хроматиды мозаичные и от их расположения на экваторе зависит, к какому полюсу они в дальнейшем отойдут. К центромерам хроматид прикрепляются нити веретена деления. Анафаза 2 (2n2с). Происходит деление двухроматидных хромосом на хроматиды и расхождение этих сестринских хроматид к противоположным полюсам клетки (при этом хроматиды становятся самостоятельными однохроматидными хромосомами), происходит третья рекомбинация генетического материала. Телофаза 2 (1n1c в каждой клетке). Хромосомы деконденсируются, образуются ядерные оболочки, разрушаются нити веретена деления, появляются ядрышки, происходит деление цитоплазмы (цитотомия) с образованием в итоге четырех гаплоидных клеток. Биологическое значение мейоза. Мейоз является центральным событием гаметогенеза у животных и спорогенеза у растений. С его помощью поддерживается постоянство хромосомного набора – после слияния гамет не происходит его удвоения. Благодаря мейозу образуются генетически различные клетки, т.к. в процессе мейоза трижды происходит перекомбинация генетического материала: за счет кроссинговера (профаза 1), за счет случайного, независимого расхождения гомологичных хромосом (анафаза 1) и за счет случайного расхождения хроматид (анафаза 2).
Вопрос 17 Взаимодействие аллельных генов. Различают несколько типов взаимодействия аллельных генов: полное доминирование, при котором рецессивный признак не проявляется, неполное доминирование, при котором у гибридов наблюдается промежуточный характер наследования, кодоминирование, в этом случае у гибридов фенотипически проявляются оба признака. Например, кодоминирование проявляется у людей с 4 группой крови. Первая группа крови у людей с аллелями i0i0, вторая — с аллелями IAIA или IAí0; третья — IВIВ или IВí0; четвертая группа имеет аллели IАIВ. Сверхдоминирование – лучшая приспособленность гетерозигот от моногибридного скрещивания (например, Аа) по сравнению с обоими типами гомозигот (АА и аа). Сверхдоминирование можно определить также как гетерозис, возникающий при моногибридном скрещивании. Наиболее известный пример взаимоотношения между нормальным (S) и мутантным (s) аллелями гена, контролирующего структуру гемоглобина у человека. Люди, гомозиготные по мутантной аллели (ss), страдают тяжёлым заболеванием крови – серповидноклеточной анемией, от которого они гибнут обычно в детском возрасте (эритроциты больного имеют серповидную форму и содержат гемоглобин, структура которого незначительно изменена в результате мутации). Однако в тропической Африке и других районах, где распространена малярия, в популяциях человека постоянно присутствуют все три генотипа SS, Ss и ss (20-40% населения гетерозиготы Ss). Оказалось, что сохранение в популяциях человека летальной (смертельной) аллели (s) обусловлено тем, что гетерозиготы (Ss) более устойчивы к малярии, чем гомозиготы по нормальному гену (SS), и, следовательно, обладают отборным преимуществом. Наследование группыABO Кодоминирование, в этом случае у гибридов проявляются оба признака. Например, кодоминирование проявляется у людей с 4 группой крови. Первая группа крови у людей с аллелями iOiO, вторая — с аллелями IAIA или IAí0; третья — IВIВ или IВí0; четвертая группа имеет аллели IАIВ. Во 2 и 3 группах полное доминирование генов IА и IВ над аллелью I0, в 4 группе в эритроцитах присутствуют продукты гена IА и IВ. Вопрос 18 Пенетрантность — частота проявления гена в фенотипе его носителей. Если у части лиц, несущих данный ген, он фенотипически не проявляется, говорят о неполной пенетрантности. Экспрессивность — степень фенотипического проявления одного и того же гена у разных лиц. Различия одного и того же признака у кровных родственников объясняются разной экспрессивностью гена, контролирующего этот признак. Разная экспрессивность встречается при большинстве моногенных болезней. Билет№ 19 В связи с тем что кариотип организма — это диплоидный набор хромосом, большинство генов в соматических клетках представлены аллельными парами. Аллелъные гены, расположенные в соответствующих участках гомологичных хромосом, взаимодействуя между собой, определяют развитие того или иного варианта соответствующего признака (см. разд. 3.6.5.2). Являясь специфической характеристикой вида, кариотип представителей разного пола различается по паре половых хромосом (см. разд. 6.1.2.1).Гомогаметный пол, имеющий две одинаковые половые хромосомы XX, диплоиден по генам этих хромосом. Гетерогаметный пол имеет одинарный набор генов Х-хромосомы (ХО) или негомологичных участков Х- и Y-хромосом. Фенотипическое проявление и наследование отдельных признаков из поколения в поколение организмов зависит от того, в каких хромосомах располагаются соответствующие гены и в каких дозах они присутствуют в генотипах отдельных особей. Различают два основных типа наследования признаков: аутосомное и сцепленное с полом (схема 6.1). |
|||||
|
|
Последнее изменение этой страницы: 2018-04-12; просмотров: 636. stydopedya.ru не претендует на авторское право материалов, которые вылажены, но предоставляет бесплатный доступ к ним. В случае нарушения авторского права или персональных данных напишите сюда... |
 Нарушения гонадогенеза при изменении числа хромосом в кариотипе Среди них известна моносомия Х (из пары хромосом присутствует только одна (Х0)) - это синдром Шерешевского-Тернера. Реже встречается трисомия Х и синдром Клайнфельтера (ХХУ, ХХХУ, ХУУ и т.д.). Люди с изменением числа половых хромосом при наличии У-хромосомы развиваются по мужскому типу. Это является следствием того, что факторы, определяющие мужской тип развития, находятся в У-хромосоме. В отличии от мутаций аутосом (все хромосомы, кроме половых), дефекты умственного развития у больных выражены не столь отчетливо, у многих оно в пределах нормы, а иногда даже выше среднего. Вместе с тем у них постоянно наблюдается нарушения развития половых органов и роста. Реже встречаются пороки развития других систем. Люди с изменением числа половых хромосом при наличии У-хромосомы развиваются по мужскому типу!! Синдром Тернера — это клиническое проявление аномалии одной из X-хромосом у женщин. Синдром Тернера в 60% случаев обусловлен моносомией X-хромосомы (кариотип 45,X), Кариотип 45,X в большей части случаев обусловлен нерасхождением половых хромосом в 1-м делении мейоза.Половой инфантилизм проявляется в пубертатном периоде: нет вторичных половых признаков, молочные железы не развиваются, вторичное оволосение отсутствует или скудное, недоразвиты половые губы, влагалище и матка. Слизистая оболочка вульвы и влагалища сухая, ее эпителиальный покров тонкий. Трисомия по X-хромосоме (47,ХХХ)взрослые больные обычно имеют нормальный женский фенотип. Синдром Клайнфельтера — это клиническое проявление полисомии по X-хромосоме у мужчин (распространенность около 1:500). Чаще всего наблюдается кариотип 47,XXY (классический вариант синдрома), но встречаются и более редкие кариотипы: 48,XXXY; 49,XXXXY; 48,XXYY; 49,XXXYY.Наружные половые органы мужского типа. Яички опущены в мошонку, плотные, очень маленькие 1,5см.(при норме 5см.). Внутренние половые органы значительно меньших размеров по сравнению с нормой. Обязательный признак заболевания - гипоплазия гонад,наблюдается гинекомастия
Нарушения гонадогенеза при изменении числа хромосом в кариотипе Среди них известна моносомия Х (из пары хромосом присутствует только одна (Х0)) - это синдром Шерешевского-Тернера. Реже встречается трисомия Х и синдром Клайнфельтера (ХХУ, ХХХУ, ХУУ и т.д.). Люди с изменением числа половых хромосом при наличии У-хромосомы развиваются по мужскому типу. Это является следствием того, что факторы, определяющие мужской тип развития, находятся в У-хромосоме. В отличии от мутаций аутосом (все хромосомы, кроме половых), дефекты умственного развития у больных выражены не столь отчетливо, у многих оно в пределах нормы, а иногда даже выше среднего. Вместе с тем у них постоянно наблюдается нарушения развития половых органов и роста. Реже встречаются пороки развития других систем. Люди с изменением числа половых хромосом при наличии У-хромосомы развиваются по мужскому типу!! Синдром Тернера — это клиническое проявление аномалии одной из X-хромосом у женщин. Синдром Тернера в 60% случаев обусловлен моносомией X-хромосомы (кариотип 45,X), Кариотип 45,X в большей части случаев обусловлен нерасхождением половых хромосом в 1-м делении мейоза.Половой инфантилизм проявляется в пубертатном периоде: нет вторичных половых признаков, молочные железы не развиваются, вторичное оволосение отсутствует или скудное, недоразвиты половые губы, влагалище и матка. Слизистая оболочка вульвы и влагалища сухая, ее эпителиальный покров тонкий. Трисомия по X-хромосоме (47,ХХХ)взрослые больные обычно имеют нормальный женский фенотип. Синдром Клайнфельтера — это клиническое проявление полисомии по X-хромосоме у мужчин (распространенность около 1:500). Чаще всего наблюдается кариотип 47,XXY (классический вариант синдрома), но встречаются и более редкие кариотипы: 48,XXXY; 49,XXXXY; 48,XXYY; 49,XXXYY.Наружные половые органы мужского типа. Яички опущены в мошонку, плотные, очень маленькие 1,5см.(при норме 5см.). Внутренние половые органы значительно меньших размеров по сравнению с нормой. Обязательный признак заболевания - гипоплазия гонад,наблюдается гинекомастия